
Myrmeleon almohadarum, Badano, Davide, Acevedo, Fernando, Pantaleoni, Roberto A. & Monserrat, Víctor J., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4196.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:75113EF0-968D-45E3-9787-9D4FCF029737 |
|
DOI |
https://doi.org/10.5281/zenodo.6084515 |
|
persistent identifier |
https://treatment.plazi.org/id/AB3C87EC-FFDC-1E48-A8BC-FF3AFDC7FF12 |
|
treatment provided by |
Plazi |
|
scientific name |
Myrmeleon almohadarum |
| status |
sp. nov. |
Myrmeleon almohadarum View in CoL sp. nov.
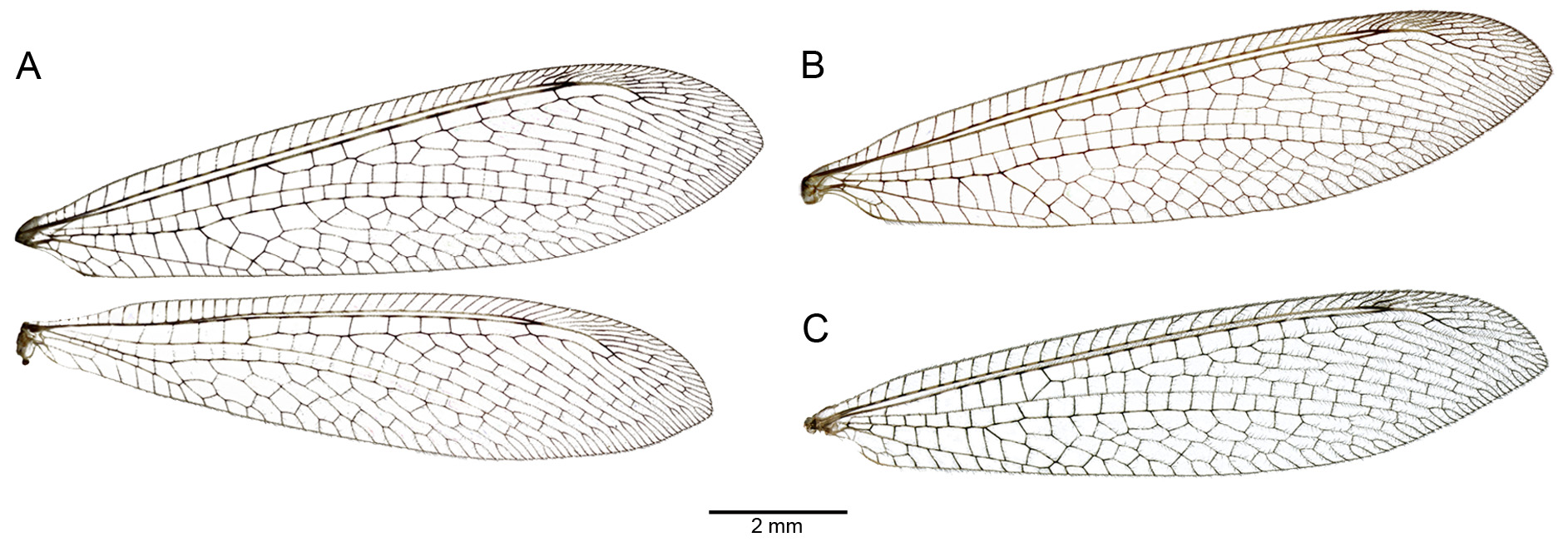
Diagnosis. Small-sized Myrmeleon with a dark brown, variegated habitus ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 ); pronotum with a diagnostic pattern ( Fig. 3 View FIGURE 3 B); wings hyaline with dark-and-pale dashed veins ( Fig. 4 View FIGURE 4 A); male hindwing with pilula axillaris; abdomen with conspicuous paler markings, creating an annulated pattern. Larva with an ochre habitus ( Fig. 7 View FIGURE 7 ), characterized by a particularly dense arrangement of digging setae on sternite IX ( Fig. 8 View FIGURE 8 ).
Description of the adult. Size. Average body length 20.43 mm (min-max 16.03–26.82); forewing male length 21.56 mm (19.20–23.33), female length 23.05 mm (19.84–26.5), ratio width/length (both sexes) 0.23; hind wing male length 19.57 mm (16.35–21.42), female length 20.81 mm (17.77–25.55), ratio width/length (both sexes) 0.22. General colouring. Dark brown with large ochre markings on thorax and abdomen ( Fig. 2 View FIGURE 2 ). Head. Vertex and occiput dark brown, with a contrasting ochre pattern ( Fig. 3 View FIGURE 3 B). Frons dark brown with paler lateral margins. Ocular rim pale. Clypeus pale with fainted median marking. Labrum pale. Maxillary palpus brown. Labial palpus pale, with the distal segment fusiform and dark brown, palpimacula elliptical. Scape brown, paler distally, pedicellum dark brown, flagellum brown. ( Fig. 3 View FIGURE 3 A). Thorax. Pronotum brown, with anterior and lateral margins pale; dorsal side with a complex pattern composed by: a narrow median pale stripe, a pair of poorly defined pale spots in the apical half and a pair of usually well contrasted pale spots in the basal half ( Fig. 3 View FIGURE 3 B). Mesonotum and metanotum brown, lateral and ventral sclerite margins ochre, particularly evident on mesoscutum, mesoscutellum, metascutum and metascutellum ( Fig. 2 View FIGURE 2 ). Legs. Coxae brown in all legs ( Fig. 2 View FIGURE 2 ). Pro- and mesothoracic legs with extensive brown markings on the femur and tibia. Metathoracic leg with brown markings on the femur and the inner face of the tibia brown ( Fig. 2 View FIGURE 2 ). Tibial spurs as long as the first tarsomere. Wings. Relatively broad with a rounded apex ( Fig. 4 View FIGURE 4 A). Membrane hyaline. Pterostigma distinct, proximally brown and distally whitish. Venation predominantly dark brown with alternating pale dashes. Radius sector of forewing with on average 5–7 (less frequently 8) crossveins, thus the cells of Rs are few and elongated. Cubital fork of forewing slightly more basal than Media posterior fork. Hind wing with on average 5 presectoral crossveins. Male hind wing equipped with pilula axillaris ( Fig. 4 View FIGURE 4 A). Abdomen. Shorter than wings ( Fig. 2 View FIGURE 2 ). Tergites brown with large dorso-proximal ochre markings. Sternites brown with ventro-proximal pale markings. Interpleural membrane brown with paler markings. Ectoproct ochre. Male terminalia as in Fig. 5 View FIGURE 5 A, B; male genitalia, complex of gonocoxites 9+11 sensu Aspöck & Aspöck 2008 (gonarcus-paramere complex) as in Fig. 6 View FIGURE 6 . Female terminalia as in Fig. 5 View FIGURE 5 C, D.
Variability. M. almohadarum sp. nov. is a relatively variable species in term of body pattern, like other congeners. Body colour varies from brown to dark brown but it is never pale ochre or blackish, like M. mariaemathildae and M. inconspicuus respectively. Some specimens are characterized by a partial fusion or fading of the pronotal pattern (e.g. specimen depicted in Fig. 2 View FIGURE 2 ). The markings on the clypeus are also relatively variable.
Description of the third instar larva. Size. Average body length 9.20 mm; head length 1.78 mm (min-max 1.67–1.87), head width 1.53 mm (1.47–1.57), mandible length 1.83 mm (1.75–1.87), ratio head width/length 0.86, ratio mandible length/head length 1.03. General colouring. Pale brown, with a dark brown pattern, ventral side of the body paler, with conspicuous dark markings ( Fig. 7 View FIGURE 7 ). Dorsal side of the head capsule with dark brown markings on the clypeo-labrum and a posterior V-shaped, fainted marking, lateral side with dark brown markings. Ventral side pale, with a pair of large dark brown spots ( Fig. 7 View FIGURE 7 ). Mandible pale brown. Some preserved Spanish larvae are characterized by the presence of a dark marking in the median section of the mandible. Legs pale brown, unspotted. Body chaetotaxy black. Head. Sub-rectangular, longer than wide. Ocular tubercle sessile. Mandible as long as the head capsule, provided with 3 equidistant teeth not abruptly differentiated in size. Interdental mandibular setae: (5)(2)(2)(1). External margin with a fringe of long setae, dorsal and ventral side almost hairless except a few short setae near the margin. Labial palpus 4-segmented. Abdomen. VIII abdominal sternite with small odontoid processes, posterior margin with stout setae. IX abdominal sternite with many ventral digging setae, unevenly arranged or irregularly aligned in rows, mostly sub-equal in size and interspersed with smaller ones ( Fig. 8 View FIGURE 8 ). Rastra sessile, each bearing 4 digging setae, of which the external pairs are longer ( Fig. 8 View FIGURE 8 ).
Examined specimens. HOLOTYPE: Spain: Cádiz: Bolonia / 30 STE 59 View Materials / 10 m / ex larva 13.VII.2015 / F. Acevedo leg., 1 ♂ in alcohol [VM] ( Fig. 1 View FIGURE 1 ) . PARATYPES. Spain: Almería: Las Casillas de Atochares, Rambla del Artal / 210 m / ex l . 20.VIII.2012 / F. Acevedo leg., 1 ♀ dry pinned [DB]; San Roque / 300 m / 2.IX.1993 / J. Ramírez leg., 1 ♀ dry pinned [VM]; Tabernas, Rambla Roja / 360 m / ex l . 16.VII.2012 / F. Acevedo leg., 1 ♂ dry pinned [DB]; Baleares: Ibiza, Las Salinas / 10 m / ex l . 3.VIII.2011 / V. J. Monserrat leg., 1 ♀ dry pinned [VM]; Cádiz: Barbate, Cerro del Pinar / 130 m / 20.VIII.1976 / V. J. Monserrat leg., 1♀ * dry pinned [VM]; Pinar de la Duquesa 26.VIII.77 E.L. / V. Monserrat 1♀ * [RAP]; Bolonia / 10 m / ex l . 13.VIII.2012 / F. Acevedo leg., 1♂ * dry pinned [VM]; Bolonia / 10 m / ex l . 25.VIII.2012 / F. Acevedo leg., 1 ♂ dry pinned [VM]; Bolonia / 10 m / ex l . 13.VII.2015 / F. Acevedo leg., 1♂ [VM]; Bolonia / 10 m / ex l . 13.VII.2015 / F. Acevedo leg., 1♂ [VM]; Bonanza / 18 m / ex l . 2.VII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [DB]; Bonanza / 18 m / ex l . 3.VII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [VM]; Bonanza / 18 m / ex l . 23.VII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [VM]; Bonanza / 18 m / ex l . 13.VIII.2012 / F. Acevedo leg., 1 ♀ * dry pinned [DB]; Cádiz, Castillo de San Sebastián / 10 m / 10.VII.1976 / V. J. Monserrat leg., 1 ♂ * dry pinned [VM]; Caños de Meca / 10 m / ex l . 23.VII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [VM]; Caños de Meca / 10 m / ex l . 4.VIII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [VM]; Chiclana / 10 m /ex l . 13.VII.2012 / F. Acevedo leg., 1 ♂ * dry pinned [DB]; Playa de Los Lances / 10 m / ex l . 13.VIII.2012 / V. J. Monserrat leg., 1 ♂ * dry pinned [DB]; Playa Los Lances / 10 m / ex l . 27.VIII.2012 / V. J. Monserrat leg., 1 ♀ * dry pinned [DB]; Puerto Santa María, La Puntilla / 10 m / 29.VI.1976 / V. J. Monserrat leg., 1 ♂ * dry pinned [VM]; Venta del Retín/ 40 m / 5.VIII.1976 / I. Reviejo leg., 1 ♀ * dry pinned [DB]; Huelva : Punta Umbría / 10 m / ex l . 16.VII.2012 / F. Acevedo leg., 1 ♀ * dry pinned [VM]; Punta Umbría / 10 m / ex l . 4.VIII.2012 / F. Acevedo leg., 1 ♀ * dry pinned [VM]; Jaén: El Centenillo / 600 m / ex l . 6.VII.1986 / V. J. Monserrat leg., 1 ♂ [VM]; El Centenillo / 600 m / ex l . 10.VII.1986 / V. J. Monserrat leg., 1 ♂ [VM]; El Centenillo / 600 m / ex l . 10.VII.1986 / V. J. Monserrat leg., 1 ♀ [VM]; El Centenillo / 600 m / ex l . 11.VII.1986 / V. J. Monserrat leg., 1 ♀ [VM]; El Centenillo / 600 m / ex l . VIII.1986 / V. J. Monserrat leg., 1 ♀ [VM]; Málaga: Estepona, Sierra Bermeja , Rio Padrón / 140 m / 9.VII.2013 V. J. Monserrat leg., 1 ♂ dry pinned [DB].
Tunisia: Tunis, Gammarth / VII.2010 / INRGREF leg. / 1♀, ex larva, in alcohol [DB] ; Tunis, Gammarth / VII.2010 / INRGREF leg. / 2♂, 2♀, ex larvae, in alcohol [RAP]; Forêt de Dar Chichou 36° 57,733N 10° 59,445E / VIII.2010 / INRGREF leg GoogleMaps . 1♂, 4♀ ex larvae, in alcohol [RAP]; Al Sawasi (=Souassi) 35° 21,953N 10° 36,689E / VIII.2010 / INRGREF leg GoogleMaps . 1♀ ex larva, in alcohol [RAP]; Tunis, Gammarth 36°55’10”N 10°17’38”E / 9.IV.2014 / R. A. Pantaleoni leg., 1 ♀, ex larva, dry pinned [DB] GoogleMaps ; Tunis, Gammarth 36°55’10”N 10°17’38”E / 9.IV.2014 / R. A. Pantaleoni leg. 1♂, 2♀, ex larvae, in alcohol [DB] GoogleMaps ; Tunis, Gammarth 36°55’10”N 10°17’38”E / 9.IV.2014 / R. A. Pantaleoni leg., 3♂, 2 ♀, ex larvae, in alcohol [RAP]. GoogleMaps
Larvae. Spain: Cádiz: Bolonia / 10 m / 24.V.2012 / F. Acevedo leg., 3 third instar larvae [VM]; Bolonia/ 10 m / 5.IV.2015 / F. Acevedo leg., 2 third instar larvae [VM]; Bonanza/ 18 m / 21.VI.2012 / F. Acevedo leg., 2 third instar larvae [VM]; Huelva: Punta Umbría / 10 m / 24.V.2012 / V. J. Monserrat leg., 3 third instar larvae [VM]; Málaga: Las Cañillas / 160 m / 25.V.2012 / F. Acevedo leg., 6 third instar larvae [VM]; Monda, Llanos de Purla / 370 m / 18.VIII.2013 / V. J. Monserrat leg., 2 third instar larvae [VM].
Tunisia: Tunis, Gammarth 36°55’10”N 10°17’38”E / 9.IV.2014 / R. A. Pantaleoni leg., 2 third instar larvae, in alcohol [DB] GoogleMaps ; Tunis, Gammarth 36°55’10”N 10°17’38”E / 9.IV.2014 / R. A. Pantaleoni leg.,over 20 larvae, in alcohol [RAP] GoogleMaps
Derivatio nominis. The name of this new species is a noun in the genitive case, referring to the Almohad dynasty, which dominated North Africa and the south of the Iberian Peninsula between the 12th and 13th centuries.
DNA taxonomy and phylogeny. The six putative species of the genus Myrmeleon , included in the analysed data set, were all supported by Bayesian posterior probabilities of 1.0 ( Fig. 10 View FIGURE 10 ). The Bayesian Poisson tree process algorithm applied on the Bayesian consensus tree also supported the existence of six species, reinforcing the morphology-based assumptions. The status of M. almohadarum as a new species was confirmed by a posterior probability of 0.97, given the input phylogeny and the bPTP model. The Bayesian phylogenetic analysis reconstructed M. almohadarum as sister to M. inconspicuus with a posterior probability of 1.0 ( Fig. 10 View FIGURE 10 ).
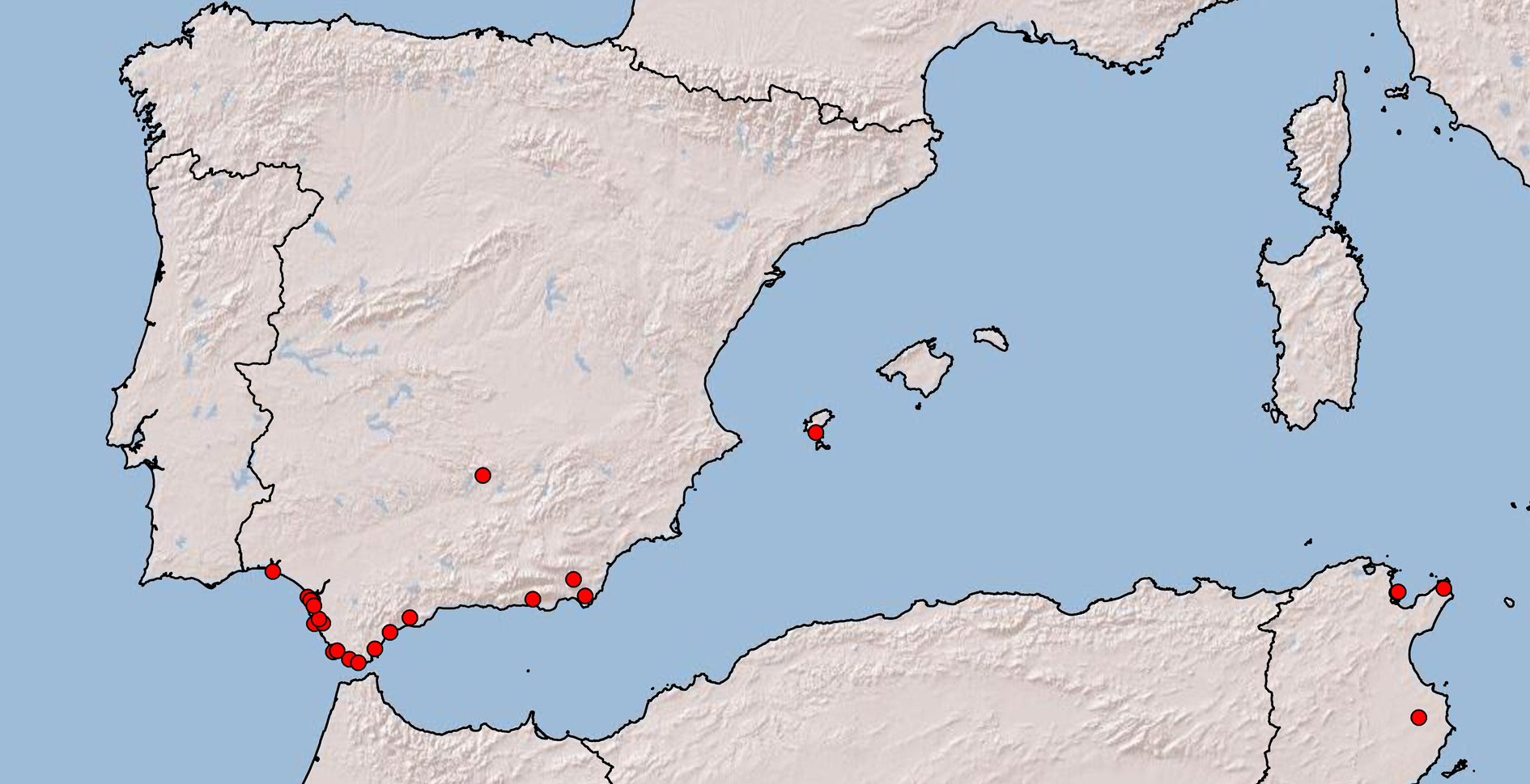
Ecological notes and distribution. In Tunisia, the pit-building larvae of M. almohadarum were collected in old coastal sand dunes with tree vegetation, including pine plantations. In Spain, besides the above mentioned environments, this species was also found in river banks and dry ephemeral riverbeds, suggesting that this antlion shares the same habitat preferences with M. inconspicuus ( Nicoli Aldini 2007; Gepp 2010; Pantaleoni et al. 2010; Badano & Pantaleoni 2014) but it is more thermophilous. M. almohadarum is presently known from relatively few localities in Spain, mostly in the southernmost part of the Iberian Peninsula (Andalusia: Huelva, Jaén, Cádiz, Málaga and Almería). An isolated record from the Balearic Islands (Ibiza) is also known ( Fig. 9 View FIGURE 9 ). At least some of the numerous records of M. inconspicuus from the Iberian Peninsula (see list in Monserrat & Acevedo 2013), especially from its southernmost part and/or the Balearic Islands, very likely belong to the new species. In North Africa, this antlion was collected in Tunisia, but it is probably widespread along the south-western Mediterranean coast, in areas influenced by the Mediterranean climate and avoiding desert environments.
Comparative notes. The Myrmeleon inconspicuus -group of species, firstly delimited by Pantaleoni et al. (2010), includes the Turano-European M. inconspicuus , the Mediterranean M. mariaemathildae and the Asiatic-E- European M. immanis Walker. These species differ from other Palaearctic congeners in a set of adult characters: forewing Cubital fork more basal than Media posterior fork ( Fig. 4 View FIGURE 4 ), male hind wing with pilula axillaris and male genitalia with a large, lamellar mediuncus ( Fig. 6 View FIGURE 6 ). Moreover, the larvae of this species-group are characterized by the IX abdominal sternite with the anterior row of digging setae composed by at least 6 bristles ( Badano & Pantaleoni 2014). M. almohadarum also fits within this species group, based on adult and larval morphology and the DNA-based phylogenetic reconstruction. The new species and M. immanis are easily distinguished by the unmistakable habitus of the latter (i.e. blackish body colour and yellowish, hyaline wings with pointed apex). M. inconspicuus is also darker than M. almohadarum and it is characterized by a blackish pronotum with a thin median line and an isolated pale spot per side ( Fig. 3 View FIGURE 3 C). On the other hand, M. mariaemathildae is a highly variable species, with distinct pale and dark morphs, though most specimens are pale ochre. The pronotum of M. mariaemathildae is usually characterized by large pale markings and paired dark areas, with a large median, pale stripe connected with a pair of posterior spots ( Fig. 3 View FIGURE 3 D), though in very dark specimens the pronotum closely resembles that of M. inconspicuus , see Pantaleoni et al. (2010) for further details. M. almohadarum is best distinguished from these closely related species due to the overall brown habitus and the dark brown pronotum with two pale markings per side connected to a median stripe (but see Variability). The new species is usually characterized by the Radius sector of forewing with about 5–7 transversal crossveins, therefore the cells of the Rs are few and comparatively long ( Fig. 4 View FIGURE 4 A). In other species (including the other members of the M. inconspicuus group) the Rs crossveins are usually 10 or more, thus the cells appear more numerous but shorter ( Fig. 4 View FIGURE 4 B, C). M. almohadarum closely resembles the African species M. caliginosus Hölzel & Ohm. The most obvious difference, besides genitalia, between these species is represented by the presence of pilula axillaris on male hind wing, always lacking in M. caliginosus . Moreover, M. caliginosus is overall darker, with pronotal spots well separated and not connected by a median line.
The larva of M. almohadarum is very similar in body pattern and relative proportions to those of M. inconspicuus and M. mariaemathildae (see Badano & Pantaleoni 2014). The larva of the new species is characterized by the IX sternite with the anterior row of digging setae comprising numerous and irregularly arranged bristles, mostly of similar size ( Fig. 8 View FIGURE 8 ). M. inconspicuus has a lower number of setae on the anterior row, while M. mariaemathildae is recognizable due to its combination of large setae disorderly interspersed with smaller ones ( Badano & Pantaleoni 2014).








| STE |
National Botanical Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |