
Monroika clarae, Bick & Armendáriz, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4966.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:2D760A10-3FE2-4CC4-A21A-46051E684131 |
|
DOI |
https://doi.org/10.5281/zenodo.4736520 |
|
persistent identifier |
https://treatment.plazi.org/id/0388879C-F561-FF87-FF14-F729FEDAF862 |
|
treatment provided by |
Plazi |
|
scientific name |
Monroika clarae |
| status |
sp. nov. |
Monroika clarae View in CoL sp. nov.
( Figs 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Manayunkia speciosa Leidy, 1858 View in CoL : Armendáriz, Paola & Rodrigues Capítulo 2011: 281–284.
Armendáriz, Spaccesi & Rodrigues Capítulo, 2012: 80–84, 89–90 ( Figs 4 View FIGURE 4 , 5A,B View FIGURE 5 ). Paola, Armendáriz, & Rodrigues Capítulo 2013: 521–529 ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ).
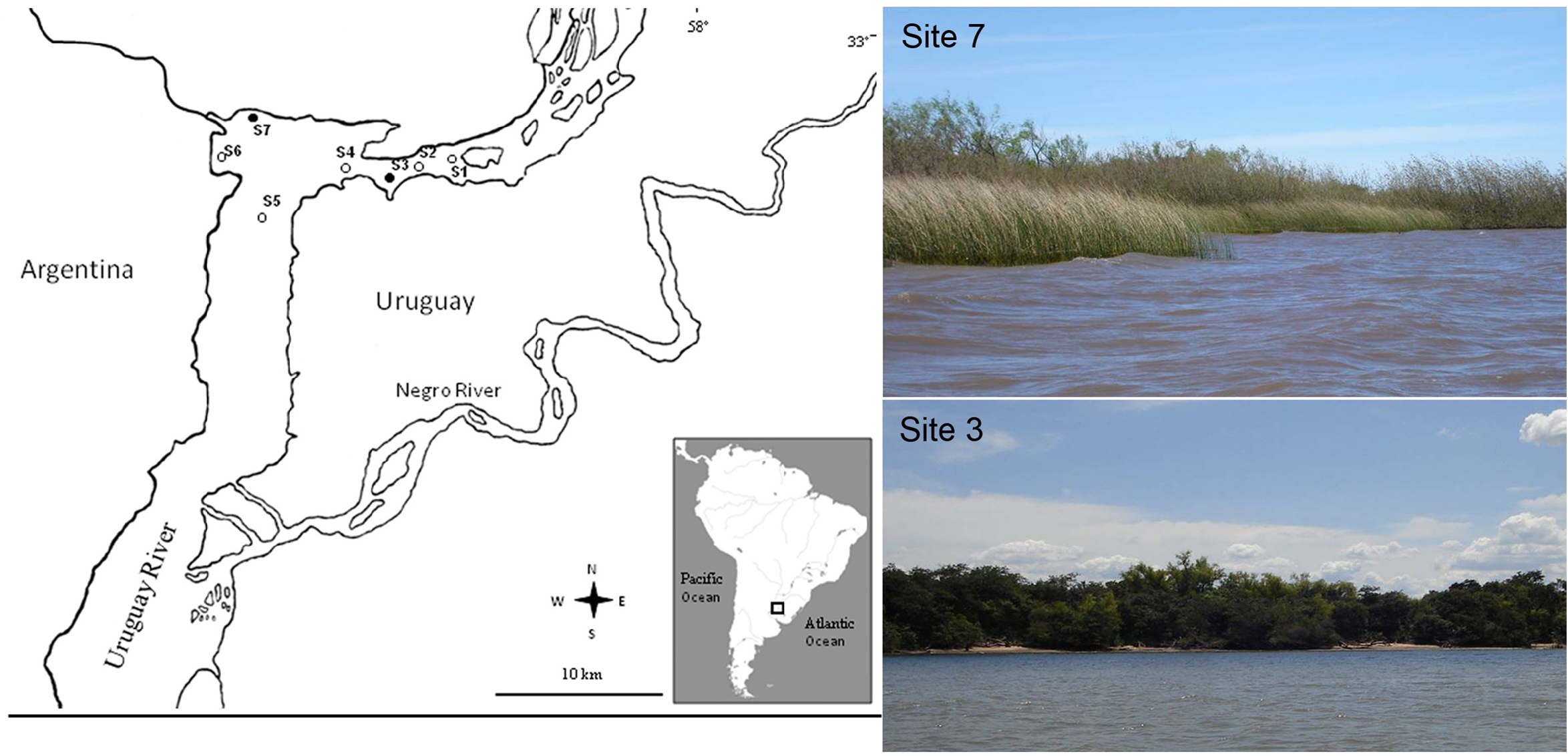
Holotype. Argentina, Lower Uruguay River, 33° 5,205’ S, 58º 22,415’ W; depth 1.5 m; September 2008 (MLP-Oi 4207). GoogleMaps
Paratypes. Argentina, Lower Uruguay River , 33° 5,205’ S, 58º 22,415’ W; depth 0.5–2.5 m, September 2008, 17 specimens (MLP-Oi 4208), 12 specimens ( ZSRO-P2651 ), 1 specimen (microscopic slides (ZSRO-2654). GoogleMaps
Additional Material. Argentina, Lower Uruguay River GoogleMaps , 33° 5,018’ S to 33º 7,441’ S, 58° 12,647’ W to 58º 25,202’ W, depth 0.5 to 9.6 m (MACN-In 37788) and (MLP-ZI-Oi 1384 (ex 6800)) (not studied here but collected from the same locality and at the same time) .
Diagnosis. Eight thoracic and two abdominal chaetigers. Pectinated radioles present, i.e. radioles asymmetrically branched. Unbranched vascularized ventral filamentous appendages present. Dorsal lips erect, rounded. Anterior margin of anterior peristomial ring developed as a low membranous collar, clearly visible only ventrally, and narrowly separated middorsally; ciliated band ventrally on the posterior margin of the anterior peristomial ring. Border between anterior and posterior peristomial rings not discernible except in the area of the ciliated band at the posterior margin of the anterior peristomial ring. Inferior thoracic notochaetae on chaetigers 2–8 narrowly hooded. Thoracic uncini of chaetigers 2–5(6?) or 2–8 (likely sexual dimorphism) with one larger tooth above main fang followed by series of smaller teeth. Abdominal uncini with one single proximal large central tooth surmounted distally by about 5 rows of equal-sized teeth, manubrium about twice as long as dentate region. Peristomial eyes black, pygidial eyes absent.
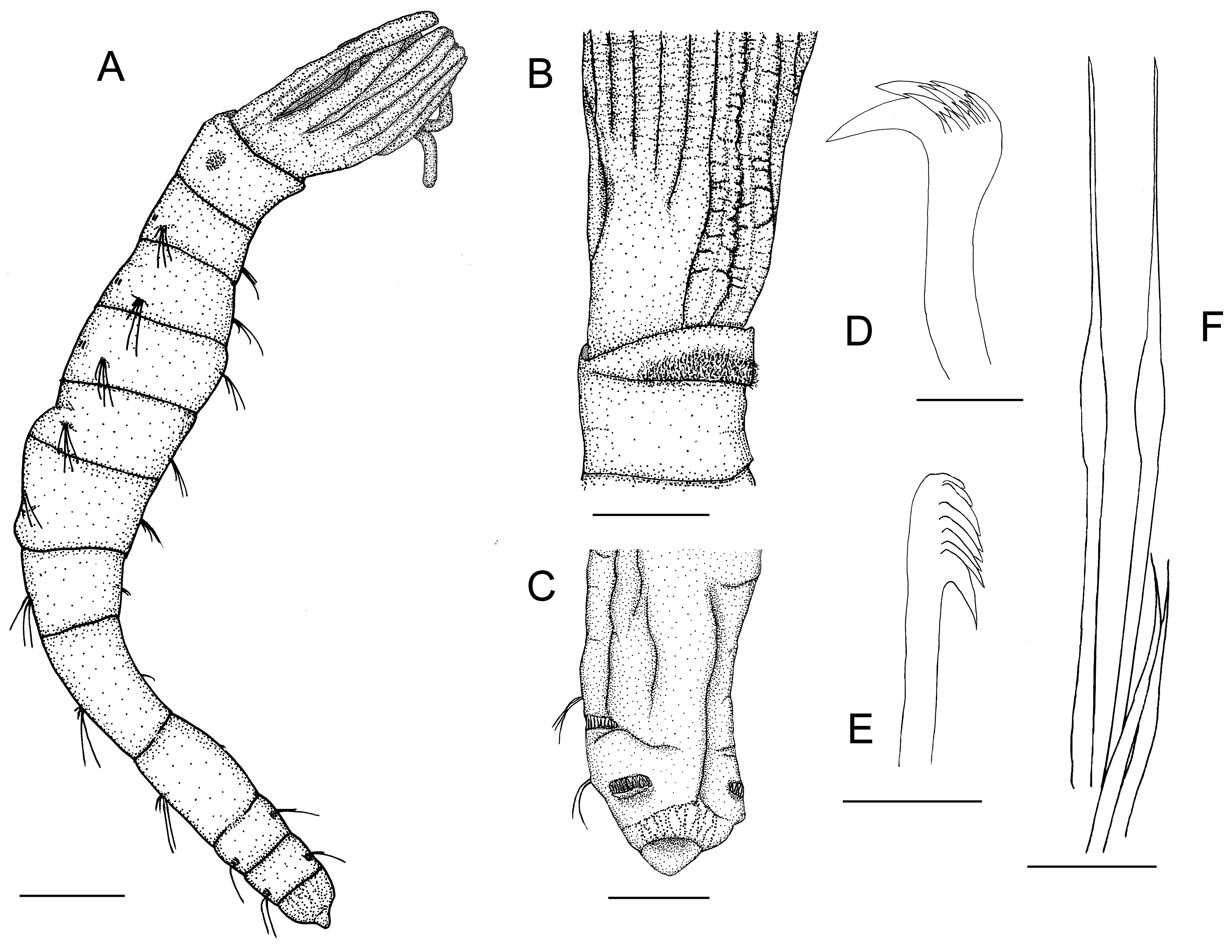
Description. Complete specimens with eight thoracic and two abdominal chaetigers ( Fig. 2A View FIGURE 2 ). Total length of the specimens, including radiolar crown, between 1.1 and 3.3 mm (holotype 2.1 mm, mean 1.8 mm), and the width between 0.08 and 0.19 mm (holotype 0.19 mm, mean 0.16 mm) ( Figs 2A View FIGURE 2 , 3A,B View FIGURE 3 ). Length of the radiolar crown between 0.19 and 0.40 mm (holotype 0.37 mm, mean 0.27 mm). Ratio between length of radiolar crown and body length between 0.14 and 0.22 (exceptionally 0.35 mm, holotype 0.21, mean 0.18 mm). Body slender, slightly tapering posteriorly ( Figs 2A,C View FIGURE 2 , 3D–F View FIGURE 3 ).
The radiolar crown is composed of three pairs of radioles and one pair of unbranched vascularized ventral filamentous appendages ( Figs 2A,B View FIGURE 2 , 3A,B View FIGURE 3 , 4A–D View FIGURE 4 ). The radiolar lobes are completely separated from one another ( Fig. 5D–H View FIGURE 5 ). The surface of the radioles and especially of the ventral filamentous appendages are wrinkled ( Figs 2B View FIGURE 2 , 4A,D View FIGURE 4 ). First the dorsal radioles branch off from the branchial lobe, then the median and then the ventral radioles ( Figs 4C View FIGURE 4 , 5A–D View FIGURE 5 ). The ventral and the median radioles are asymmetrical branched or pectinated, the dorsal radioles are unbranched ( Figs 4B,C View FIGURE 4 , 5A–D View FIGURE 5 ). The ventral radioles have a total of 5 (left radiole) and 7 (right radiole), and the median radioles a total of 4 (left radiole) and 5 (right radiole) branches (only one specimen examined) ( Fig. 4C View FIGURE 4 ). There are no morphological differences in the structure of the various branches of the radiolar crown, except the ventral filamentous appendages ( Figs 4B View FIGURE 4 , 5A,B View FIGURE 5 ). These appendages and almost all branches of the radioles end at about the same height ( Figs 2A View FIGURE 2 , 3A,B View FIGURE 3 , 4A,C,D View FIGURE 4 ). All branches of the radioles and the vascularized ventral filamentous appendages are rectangular to square in cross-section and have an extension of about 15–20 x 18–28 µm and of about 30–35 x 32–37 µm, respectively ( Figs 4B View FIGURE 4 , 5A–F View FIGURE 5 ). There is a clearly visible blood vessel in the ventral filamentous appendages ( Fig. 5A–F View FIGURE 5 ). The epidermis of the radioles consists adorally of ciliated and aborally of non-ciliated epithelial cells. The ciliated cells constitute the shallow or even convex food groove of the radioles ( Figs 3C View FIGURE 3 , 5A–D View FIGURE 5 ). The lateral cilia of the food groove are clearly longer than the median cilia ( Fig. 3C View FIGURE 3 ). The center of all branches of the radioles is occupied by a large cavity bordered by a narrow band of cells. This is followed by a thin layer of an extracellular matrix ( Fig. 5A–C View FIGURE 5 ). The dorsal lips are developed as triangular lobes, rounded at the upper margin ( Figs 4C View FIGURE 4 , 5E View FIGURE 5 ). They have a length of about 20 to 30 µm. Ventral lips or lip-like processes are absent.
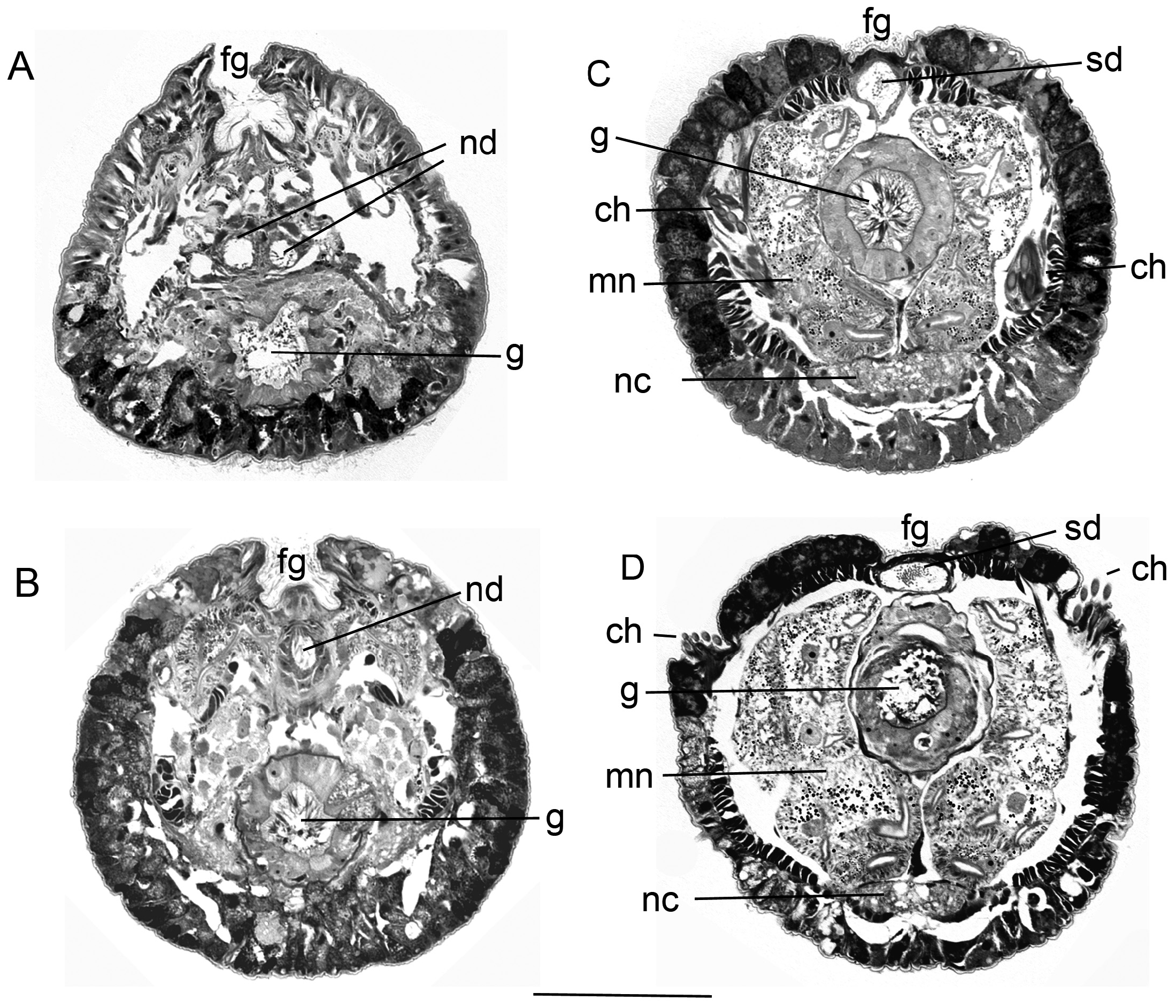
The peristomium consists of an anterior and a posterior ring. The anterior ring is distinctly shorter than the posterior ring ( Fig. 2B View FIGURE 2 ). The anterior margin of the anterior peristomial ring is developed as a low membranous collar, clearly visible only ventrally, and narrowly separated middorsally ( Figs 3A,B View FIGURE 3 , 4A,D View FIGURE 4 , 5G,H View FIGURE 5 ). A ciliated band is present ventrally on the posterior margin of the anterior peristomial ring ( Figs 2B View FIGURE 2 , 4A View FIGURE 4 , 5H View FIGURE 5 ). The border between the anterior and posterior peristomial rings is not discernible except in the area of the ciliated band. One pair of black peristomial eyes is present. The spermathecae of the females are also distinctly pigmented. They are located in the anterior peristomial ring, and could be misinterpreted as a second pair of eyes.
The faecal groove is dorsally deeply sunken in the peristomial region, and becomes considerably lower in the area of the first and the following chaetigers ( Figs 3A View FIGURE 3 , 4D View FIGURE 4 , 6A–D View FIGURE 6 ). It shifts from the dorsal midline to the ventral midline at the border between thorax and abdomen ( Fig. 3D View FIGURE 3 ).
The metanephridia are located in the peristomium and the first chaetiger ( Fig. 6 View FIGURE 6 ). The nephridial duct is paired first and then becomes unpaired. The nephroporus is located between the basal radiolar lobes ( Fig. 5H View FIGURE 5 ). In males, the sperm duct is located directly under the faecal groove ( Fig. 6C,D View FIGURE 6 ).
The first chaetiger is slightly shorter than the peristomium and the second chaetiger, chaetigers 3–6 or 7 are each successively longer, chaetigers 6 or 7 and 8 are about the same length. First chaetigers wider than long, and last 2 or 3 thoracic chaetigers distinctly longer than wide ( Figs 2A View FIGURE 2 , 3A,B View FIGURE 3 ). Abdominal chaetigers short, abdomen in total shorter than the last thoracic chaetiger. Pygidium of about same length as chaetiger 10, terminating as rounded lobe ( Figs 2A,C View FIGURE 2 , 3D–F View FIGURE 3 ). Pygidial eyes are absent. The border between chaetigers is sometimes indistinct, especially on the posterior chaetigers ( Fig. 3D–F View FIGURE 3 ).
First chaetiger with about 1–3 short, and 2–4 elongate, narrowly hooded notochaetae (not discernible in the holotype). Neuropodial uncini are absent ( Fig. 3A,B View FIGURE 3 ). Notopodia of chaetigers 2–8 superiorly with about 3–5 elongate and inferiorly with 2–3 short narrowly hooded chaetae (holotype with 3-4 elongate and 2–3 short chaetae) ( Figs 2F View FIGURE 2 , 3G,H View FIGURE 3 ). Neuropodia of chaetigers 2–6 or 2–8 with 2-4 (rarely 5) uncini with one large tooth above main fang followed by series of smaller teeth (holotype with 2–3 uncini). The single large tooth is slightly offset in relation to the main fang ( Figs 2D View FIGURE 2 , 3I,J View FIGURE 3 ). In a number of specimens, transitional uncini were found on the chaetigers 7 and 8 that were distinctly different from the thoracic uncini on chaetigers 2–6. There are usually 3 transitional uncini at the position of the regular neuropodial uncini. A clear main fang is not present here. Instead, there are a multitude of rows of small, equal-sized teeth ( Fig. 3K,L View FIGURE 3 ). Abdominal neuropodia with 1-3 elongate narrowly hooded chaetae (holotype with 3 chaetae on chaetiger 9 and 2 on chaetiger 10), and abdominal notopodia with 10–17 uncini on chaetiger 9, and 5–15 uncini on chaetiger 10 (holotype with 12(?) and 8 uncini, respectively). Abdominal uncini with one single large central tooth proximally surmounted distally by about 5–6 rows of equal-sized teeth, about 3–5 teeth per row ( Figs 2E View FIGURE 2 , 3N–P View FIGURE 3 ). The manubrium is about twice as long as the dentate region.
Fixed specimens have no conspicuous pigmentation.
Remarks. These specimens have characters that allow a clear assignment of this new species to the genus Monroika . These are mainly the presence of asymmetrically branched radioles, unbranched vascularized ventral filamentous appendages and two abdominal chaetigers. To this genus belongs only one species, M. africana . Monroika clarae sp. nov. differs clearly from M. africana by the absence of pseudospatulate chaetae on thoracic chaetigers. They are present on chaetigers 2 or 3–5 in M. africana ( Fitzhugh 1992, Jones 1974). Furthermore, M. africana possesses 3–5 radiolar branches and 18–41 abdominal uncini whereas M. clarae sp. nov. has 4–7 radiolar branches and 5–17 uncini on abdominal chaetigers.
A total of 360 specimens were found in the Lower Uruguay River but only 35 individuals were available for this study. Fifteen specimens were studied earlier by Paola et al. (2013). However, the presence of 3 abdominal chaetigers was not correctly described in this paper, because their Fig. 3 View FIGURE 3 clearly shows that only 2 chaetigers are present. Their specimens were slightly larger than those we examined: mean total length was 2.3 mm compared to 1.8 mm, mean width 0.24 rather than 0.16 mm, and mean length of radiolar crown 0.37 mm instead of 0.27 mm.
Transitional uncini on chaetigers 7 and 8 were found in some specimens of M. clarae sp. nov. ( Fig. 3K,L View FIGURE 3 ). A main fang is absent on these uncini, but a multitude of rows of small, equal-sized teeth are present. However, these uncini were not found in all specimens, so that a sexual dimorphism might be present (see Discussion). Furthermore, it is not certain whether these uncini start at chaetiger 7 or already at chaetiger 6. The corresponding uncini at chaetiger 6 were broken off at this chaetiger in all examined individuals.
Etymology. This species is named after Clara, the granddaughter of the first author (AB), who will start studying biochemistry this year.
Geographic distribution. Known only from the type locality, Lower Uruguay River, Argentina.
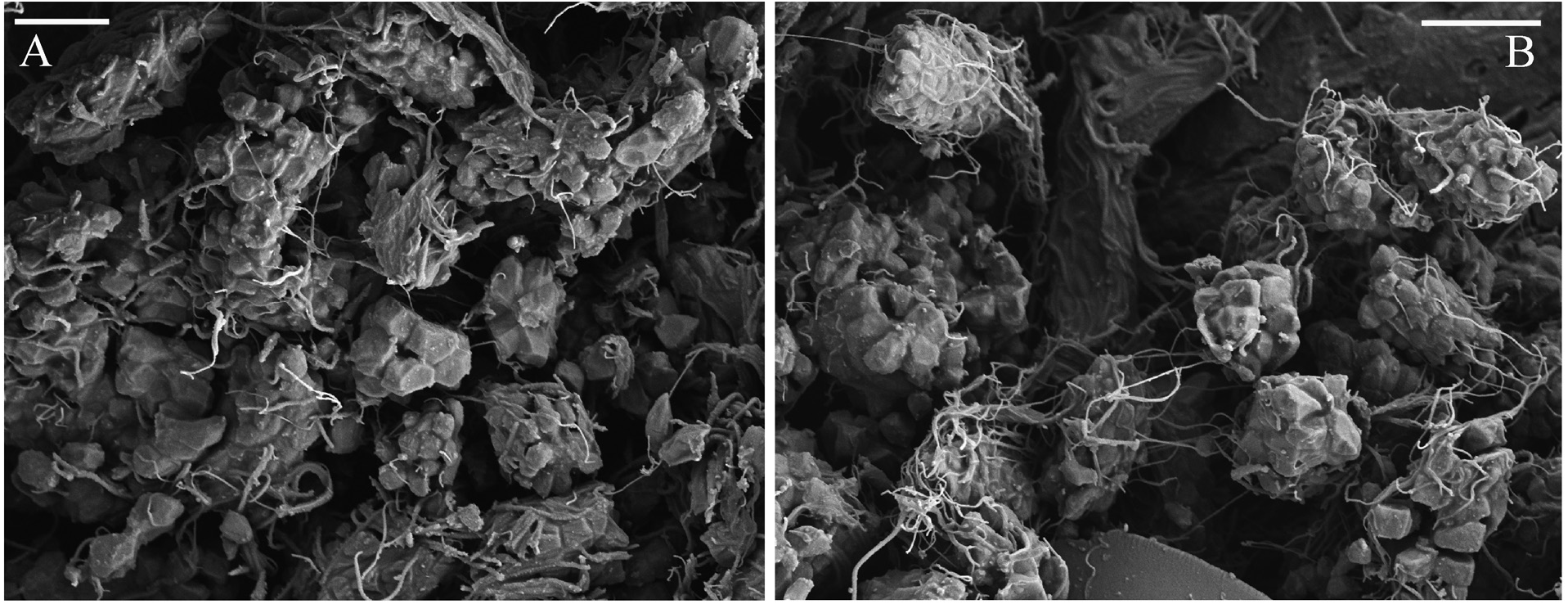
Biology. In the material used for this study and consisting of 35 individuals there was one female in a tube with three eggs. These were 160 x 118 µm, 163 x 109 µm, and 170 x 112 µm, and on average 164 x 113 µm in size. In one damaged male, SEM examinations revealed large clusters of gametes at various stages of development in the posterior thoracic chaetigers ( Fig. 7 View FIGURE 7 ).
Ecology. This species was found mainly in calm areas with aquatic vegetation and high organic matter content. The highest abundance was 2,890 ind. m-2, and average abundances were recorded at site S7 with 523 ind. m- 2 in November 2008, and at site S3 with 517 ind. m- 2 in June 2008 ( Fig. 1 View FIGURE 1 ). The main companion taxa were the mollusks Limnoperna fortunei , Corbicula fluminea (both are invasive species in America), Heleobia sp. , Potamolithus sp. , and also oligochaetes, ostracods and insects ( Chironomidae , Elmidae and Caenis sp. ). For more details see Armendáriz et al. (2011, 2012).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Monroika clarae
| Bick, Andreas & Armendáriz, Laura 2021 |
Manayunkia speciosa
| Armendariz, L. C. & Paola, A. & Rodrigues Capitulo, A. 2011: 281 |