
Miconia neocoronata Gamba & Almeda, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.179.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5156347 |
|
persistent identifier |
https://treatment.plazi.org/id/03C887CB-FB0E-FFE6-FACB-EB3EFE9D5DBD |
|
treatment provided by |
Felipe |
|
scientific name |
Miconia neocoronata Gamba & Almeda |
| status |
nom. nov. |
21. Miconia neocoronata Gamba & Almeda View in CoL , nom. nov. Basionym: Clidemia coronata Gleason (1939a: 114–115) . Type: COSTA RICA. Bords du Río Tuis, July 1893, Pittier 8080 (holotype: US-internet image!; isotypes: BR-3 sheets-internet images!). Nec Miconia coronata (Bonpland) de Candolle (1828: 187) .
Small shrub, rarely epiphytic, (0.5–)1–2(–2.75) m tall, loosely and irregularly branched. Upper internodes [1.75–4.25 cm long] and cauline nodes terete, nodal line absent. Indumentum on branchlets, petioles, primary, secondary, tertiary leaf veins abaxially, inflorescence axes, bracts, bracteoles, and pedicels (when present) densely to copiously composed of brownish sessile or thinly stipitate dendritic trichomes 0.2–0. 5(–1.9) mm long with short axes and few-moderate number of terete arms, rarely sparsely intermixed with caducous elongate smooth trichomes 1–1.5 mm long. Leaves of each pair somewhat anisophyllous in size; the petiole 0.5–3.9 cm long, canaliculate adaxially and shallowly grooved abaxially; larger blades 9–16 × 5–9.5 cm, smaller blades 3.5–10 × 2.1–5.5 cm, ovate to elliptic-ovate, the base rounded to obtuse, the margin ciliate and repand-entire, the apex shortacuminate, chartaceous; mature leaves with adaxial surface, primary, secondary, tertiary and higher order veins glabrous; abaxial surface essentially glabrous except for a few dendritic trichomes and resinous glands on the venules, the dendritic trichomes on the secondary and tertiary veins sparsely intermixed with caducous resinous slightly furrowed more or less stalked glands ca. 0.08 mm long, the higher order veins moderately beset with the same type of resinous glands; 5- or 7-plinerved, including the tenuous marginals, innermost pair of secondary veins diverging slightly to notably asymmetrically from the primary vein 0.25–0.35 cm above the base, areolae 0.5–0.75 mm, adaxially the primary and secondary veins slightly impressed to flat, the tertiary and higher order veins flat, abaxially the primary and secondary veins elevated and terete, the tertiary and higher order veins slightly elevated. Inflorescences a pseudolateral group of few-flowered modified dichasia 1.5–4 cm long, sessile, commonly with three or more paracladia arising from the base, borne on the upper foliar nodes, the rachis together with bracts and bracteoles pale magenta; bracts 0.65–0.75 × ca. 0.25 mm, spatulate-oblong, aristate at the apex, decurrent at the base, sometimes with a swollen glabrous structure at the base abaxially, both surfaces glabrescent with age, persistent in flower and tardily deciduous in fruit; bracteoles 0.65–0.95 × 0.21–0.49 mm, sessile, spatulate-oblong or triangular, the apex aristate-ciliate, the base decurrent, with one visible central vein, both surfaces glabrescent with age, persistent in fruit. Flowers 5-merous, sessile or on pedicels <0.4 mm long. Hypanthia at anthesis 3–3.2 × 0.95–1.25 mm, free portion of hypanthium 0.8–1 mm long, urceolate and constricted distally into a cylindric neck, bluntly 10-ribbed, bright pink, copiously and caducously resinous with slightly furrowed more or less stalked glands ca. 0.05 mm long, sparsely intermixed with the general and caducous dendritic trichomes, ridged on the inner surface, glabrous, the torus densely to moderately ciliolate adaxially, rarely glabrous. Calyx open in bud and deciduous on fruiting hypanthia; tube 0.15–0.3 mm long, adaxially sparsely ciliolate to glabrescent, abaxially
SYSTEMATICS OF THE OCTOPLEURA CLADE OF MICONIA
Phytotaxa 179 (1) © 2014 Magnolia Press 95 sparsely resinous glandular with the same type of glands as the hypanthium; lobes 0.15–0.45 × 0.65–0.85 mm, rounded-triangular, the margin entire to vaguely undulate, the apex subacute, sparsely and caducously papillose on both surfaces; exterior calyx teeth ca. 0.5 mm long, subulate, inserted at the base of the calyx lobes, equaling or occasionally exceeding the lobes in length, caducously resinous-glandular on both surfaces. Petals 0.75–1.15 × 0.65–0.85 mm, obovate and somewhat asymmetrical, the margin entire to sinuate, the apex rounded-obtuse, rather asymmetric, cream to white, densely papillose on both surfaces, reflexed at anthesis. Stamens 10; filaments 0.95–1.25 × 0.25 mm, whitish, glabrous; anther thecae ca. 2 × 0.33–0.34 mm, linear-oblong and subulate, truncateacuminate at the apex, opening by one dorsally inclined pore 0.11–0.13 mm in diameter, pale yellow to yellow at anthesis; connective yellow, its prolongation and appendage 0.35–0.45 mm long, the appendage oblong-spatulate to somewhat deltoid, obtuse to truncate at the apex, minutely and caducously glandular, the glands sessile and rounded. Ovary 5-locular, completely inferior, 2–2.2 mm long at anthesis, the apical collar absent, the apex 0.13–0.2 mm in diameter, slightly depressed, caducously glandular-puberulent; style ca. 5 mm long, parallel-sided (i.e. terete), white, glabrous; stigma truncate to expanded truncate at anthesis. Berries 3.1–4.1 × 3–3.9 mm when dry, globose, initially bright pink but ripening bright purple-black, the hypanthium indumentum subpersistent at maturity. Seeds 0.3–0.45 × 0.16–0.18 mm, ovoid, angled, light-brown; lateral and antiraphal symmetrical planes ovate, the highest point near the central part of the seed; raphal zone suboblong, ca. 20% larger than the corpus of the seed, extending along its entire length, ventrally and longitudinally expanded, dark-brown; individual cells elongate, anticlinal boundaries channeled, undulate, with Ω- and U-type patterns; periclinal walls convex, lowdomed to nearly flat, microrelief punctate.
Additional specimens studied:— COLOMBIA. Chocó: (Bahía Solano), Ciudad Mutis, Quebrada Seca , at waterfall near by, 29 m, 6.2°N, - 77.4°W, 6 February 2012, Almeda et al. 10470 ( CAS, CHOCO, COL) GoogleMaps . COSTA RICA. Cartago: In forest in mountains above the Río Pacuare, near Platanillo , 800 m, 3 May 1956, Williams 19517 ( EAP, US) ; Pendiente muy empinada que va desde los edificios del Instituto Interamericano de Ciencias Agrícolas de Turrialba hacia el Río Reventazón , 600 m, 19 September 1964, Jiménez 2380 ( CR, NY); (Turrialba) , Forested slope leading down to the Río Reventazón behind main building of CATIE, 9°53.5’N, 83°39’W, 560–600 m, 30 July 1985, Grayum & Hammel 5755 ( CAS, MO); (Turrialba) GoogleMaps , Across the Río Reventazón from Interamerican Institute in Turrialba , 610 m, 21 July 1947, DeWolf 369 ( US) ; Bosque húmedo y empinado entre el Inst, de Turrialba y el Río Reventazón. , 600 m, 12 July 1965, Jiménez 3276 ( CR, F, NY, US) . San José: (Tarrazu), Estribaciones del Cerro Diamante , 9°32’30"N, 84°1’20"W, 500–600 m, 23 June 1998, Estrada et al. 1636 ( CAS, CR); (Pérez Zeledón) GoogleMaps , Basin of El General , 9.1°N, - 83.29°W, 675–900 m, May 1940, Skutch 4948 ( MO, US) GoogleMaps .
Illustration:— None found.
Common names and documented uses:— None recorded.
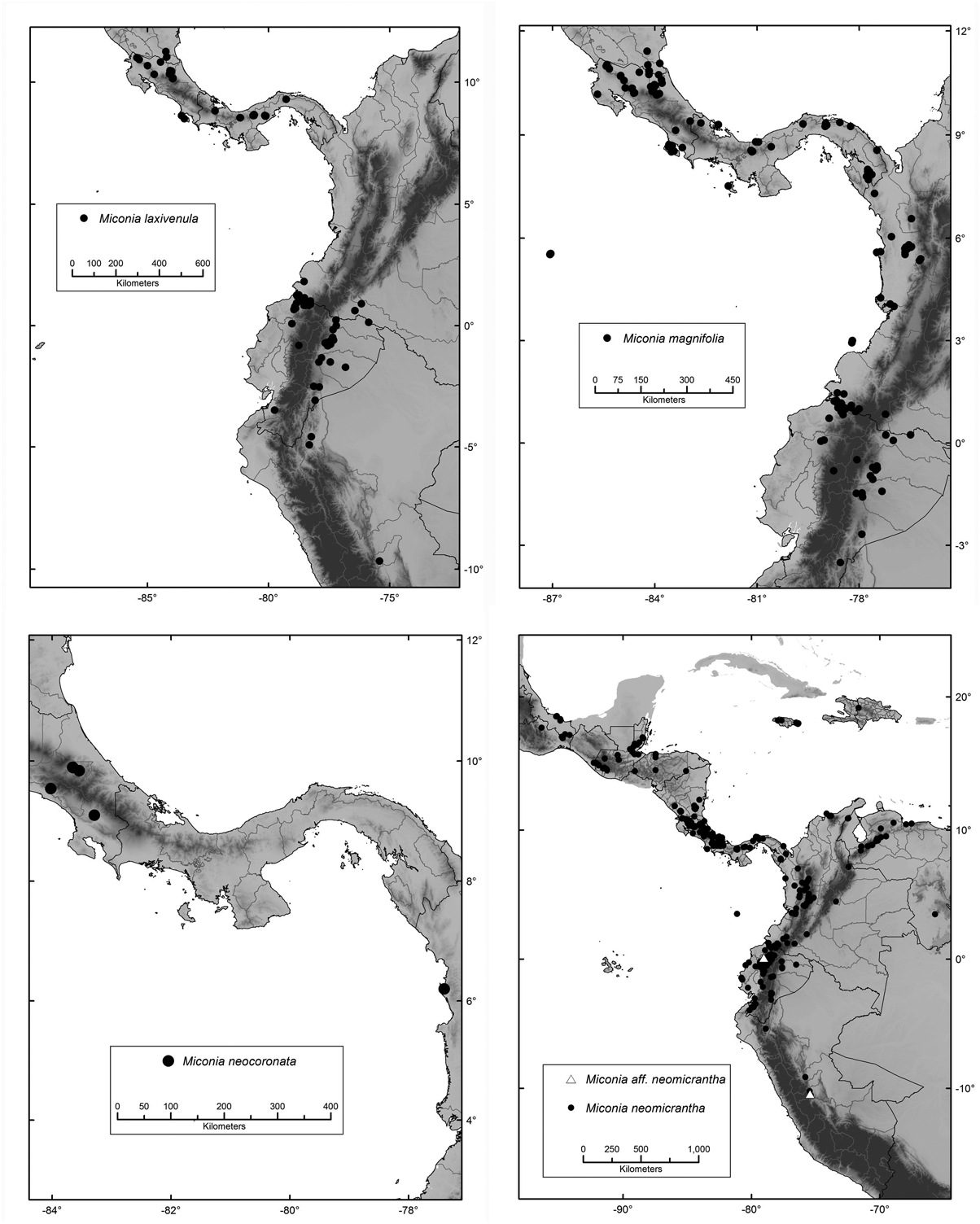
Habitat, distribution and ecology:— This is a rare species known from few collections in the low-elevation forested slopes of primary rain and riparian forests, commonly near rivers, in Costa Rica at 560–900 m. Although M. neocoronata has long been considered to be endemic in Costa Rica, it was recently collected in Colombia, from the tropical wet forest in the department of Chocó at 29 m ( Almeda et al. 10470, CAS!, CHOCO, COL). This occurrence suggests that the geographic and elevational range of this species is more extensive than expected. It probably occurs in suitable habitats in intervening areas of Panama ( Fig. 15 View FIGURE 15 ) .
Phenology:— Collected in flower and fruit in February, from May through July, and in September.
Etymology:— The specific epithet comes from the Greek word neo (new), in reference to this new combination, and from the Latin word coron (a crown), that probably refers to the minute pubescence in the ovary apex. Gleason did not explain the designation of the name coronata in the protologue.
Discussion:— Miconia neocoronata is distinct from its closest relatives by elimination of characters, particularly indumentum details. It differs from M. atropurpurea , its sister species, in hypanthial indumentum; in M. atropurpurea it consists of elongate smooth pinkish trichomes, and in M. neocoronata by the slightly furrowed more or less stalked resinous glands. Vegetatively and in floral characters M. neocoronata is very similar to M. quinquenervia . See Almeda (2004) for a thorough discussion of differences. Almeda (2004) commented on the slight difference in their connective appendages; although they are similar in shape and both are glandular, in M. neocoronata the edge is entire (vs. gland-edged). These two species are also close to M. reitziana , which shares the rusty-asperous vegetative indumentum and the resinous-glandular hypanthium, but lacks the decurrent leaf bases present in M. quinquenervia and its venation is prevailingly 7-plinerved (vs. 9-plinerved). Miconia quinquenervia is also distinct in its inflorescence architecture (dichasial dithyrsoid vs. groups of modified dichasia). The rusty-
96 Phytotaxa 179 (1) © 2014 Magnolia Press
GAMBA & ALMEDA
asperous trichomes in M. reitziana are almost concealed by the typically dense pink or red indumentum, rarely present throughout and completely absent on the foliar surfaces except for the margins in M. neocoronata . Miconia neocoronata has unique petal morphology, being obovate and somewhat asymmetrical (vs. more oblong in M. quinquenervia and M. reitziana ), and its hypanthium is constricted above the ovary into a neck (vs. subcylindric to campanulate in M. reitziana and urceolate to campanulate in M. quinquenervia ).
Conservation status:— Endangered EN B2ab(iii). This species is rare and not known from any protected areas in CostaRica.
| CAS |
California Academy of Sciences |
| CHOCO |
Universidad Tecnológica del Chocó |
| COL |
Universidad Nacional de Colombia |
| EAP |
Escuela Agrícola Panamericana |
| US |
University of Stellenbosch |
| CR |
Museo Nacional de Costa Rica |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| MO |
Missouri Botanical Garden |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |