
Macrobiotus derkai, Degma, Peter, Michalczyk, Łukasz & Kaczmarek, Łukasz, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181336 |
|
DOI |
https://doi.org/10.5281/zenodo.5625776 |
|
persistent identifier |
https://treatment.plazi.org/id/039887DC-4F1C-C755-4CF8-DB0DFED34ED1 |
|
treatment provided by |
Plazi |
|
scientific name |
Macrobiotus derkai |
| status |
sp. nov. |
Macrobiotus derkai sp. nov.
Figs. 1–74 View FIGURES 1 – 2 View FIGURES 3 – 5 View FIGURES 6 – 9 View FIGURES 10 – 11 View FIGURES 12 – 26 View FIGURES 27 – 29 View FIGURES 30 – 34 View FIGURES 35 – 38 View FIGURES 39 – 47 View FIGURES 48 – 51 View FIGURES 52 – 66 View FIGURES 67 – 72 View FIGURE 73 View FIGURE 74 , Tables 1–4
Type locality: North-eastern Colombia, Cordillera Oriental, Sierra Nevada del Cocuy National Park, near Laguna de la Plaza (mountain lake), 06°26’ N, 72°15’ W, ca. 4,300 m a.s.l.
Type material: Holotype – sex unidentified, leg. T. Derka, 11 January 2006, moss from rock; mounted in Hoyer’s medium. The microscope slide with the holotype (350/20) is deposited in the Department of Zoology, Comenius University, Bratislava, Slovakia.
Paratypes: 50 animals (including juveniles) and 88 eggs (including 7 egg shells, 39 eggs with visible embryos and 5 eggs with hatching juveniles) from the same sample as the holotype on 35 microscope slides (350/1, 350/3–9, 350/11–19, 350/21–38; all in Hoyer’s medium). Two slides are deposited in the collection of Łukasz Kaczmarek (Department of Animal Taxonomy and Ecology, A. Mickiewicz University, Poznań, Poland), two slides in the collection of Łukasz Michalczyk and two slides in the collection of Maria Grazia Binda & Giovanni Pilato (Department of Animal Biology ”Marcello La Greca”, University of Catania, Italy). Remaining slides are in the same depository as the holotype.
The 11 paratypes (5 adults and 6 eggs) prepared for SEM are not deposited in any collection.
Etymology: The new species is named after Tomáš Derka (Comenius University, Bratislava), who kindly collected the sample for us.
Diagnosis: Medium sized Macrobiotus with eyes. Cuticle smooth, without pores. Granulation on all legs present. All three bands of teeth in the oral cavity fully developed. Ventro-median tooth in the third band of the oral cavity armature most often subdivided into several teeth. Pharynx with three macroplacoids and without a microplacoid. The second macroplacoid is the shortest and it is placed closer to the first than to the third macroplacoid. Claws of the hufelandi - type, with smooth lunules and long, protruding accessory points. Eggs with short conical processes with a single short tip. Terminal parts of egg processes and their tips are covered by fine irregular tubercles. Egg areolation of the huziori - type (mostly oval areolae usually transversally divided into two shallow areolae and each of them usually deepens into two smaller areolae; spaces between neighboring areolae are usually broader than their widths).
Description: Adults and juveniles (measurements of the holotype; other measurements and their pt ratios are provided in Tables 1–2): The body of median length, transparent/white ( Figs. 1–5 View FIGURES 1 – 2 View FIGURES 3 – 5 ). Posterior eyes present (poorly visible or not visible in specimens fixed in the Hoyer’s medium and not visible in hatching juveniles). Cuticle smooth in LM, however wrinkled in SEM ( Figs. 8–9 View FIGURES 6 – 9 ), without pores but with fine regular granulation on external surface of all legs (0.1–0.3 μm; LM), better visible on hind legs ( Fig. 34 View FIGURES 30 – 34 ). Granulation may not be visible in just hatched specimens. In SEM, the complex structure of these granules is detectable: each granule consists of small cones (2–8 cones on each granule). On the dorsal side of the head a round cuticular depression is visible in SEM ( Figs. 6–7 View FIGURES 6 – 9 ).
Bucco-pharyngeal apparatus of the Macrobiotus type ( Figs. 10–11 View FIGURES 10 – 11 ). Mouth opening antero-ventral, surrounded by a ring of 10 well developed peribuccal lamellae. The oral cavity armature very well developed, consisting of three bands of teeth ( Figs. 12–26 View FIGURES 12 – 26 ).
The first band of teeth covers the posterior part of peribuccal lamellae. It is composed of small (0.2–0.6 μm, mostly 0.3–0.4 μm in diameter) dense teeth in the shape of round or slightly oval granules/cones (spaces between teeth are usually smaller than teeth themselves) (LM). This band is continuous and looks the same on all oral cavity walls, usually consisting of approximately 6–7 irregular rows of teeth (partly visible on Figs. 20 and 23 View FIGURES 12 – 26 ).
The second band of teeth is placed in the posterior portion of the oral cavity just before the third band of teeth. This band is continuous and arranged in two sub-bands of teeth: an anterior and a posterior band of teeth. The teeth in the anterior band are in the shape of small ridges parallel to the main axis of the buccal tube and teeth in the posterior band (situated just before the third band of teeth) are in the shape of granules/cones. The teeth in the anterior band are regular, however often joined two by two (sometimes three together) mainly in their posterior parts. Joined teeth are V- or W-shaped. The teeth of this row are relatively wide (0.3–0.4 μm) and their lengths are not equal; the shortest are those on the lateral walls of the oral cavity and the dorsal ones are slightly longer (0.7–1.2 μm) than the ventral ones (0.7–0.8 μm). The posterior teeth of the second band of teeth are generally similar to those of the first band in their shape and size but are more irregularly distributed (less dense on the lateral walls of the oral cavity). The band of these small teeth is wider ventrally than dorsally ( Figs. 19–20 View FIGURES 12 – 26 ).
The third band of teeth is not continuous and consists of two series (dorsal and ventral) of large, transverse ridge-shaped and round or oval teeth. The band is positioned in the rear of the oral cavity just behind the second band of teeth and just before the buccal tube opening. The three dorsal teeth are in the shape of ridges and they are situated more anteriorly than the ventral teeth of this band. The dorso-median tooth is transversally oval or rectangular with rounded edges (length and width are 1.8 x 1.0 μm) and it is usually distinctly separated from the dorso-lateral teeth, which are in the shape of long and thin ridges (lengths and widths: 3.6–4.3 x 0.8–0.9 μm). The ventro-median tooth is usually fragmented into 2–4 round or oval teeth ( Figs. 12–28 View FIGURES 12 – 26 View FIGURES 27 – 29 ), most frequently two ventro-median teeth are present (two in the holotype with the lengths and widths of 0.9– 1.0 x 0.6–0.7 μm). The ventro-lateral teeth are wing-shaped, thicker in their median portions (lengths and widths: 3.0–3.1 x 0.7–0.9 μm). The dorso-lateral teeth are longer and slightly closer to each other than the ventro-lateral ones ( Figs. 12–18, 22–26 View FIGURES 12 – 26 ).
Buccal tube relatively wide (Tables 1–2) with an anterior bend ( Fig. 11 View FIGURES 10 – 11 ). Stylet supports inserted on the buccal tube farther down than three quarters of its length (Table 2). Pharyngeal bulb slightly oval (72.4 μm x 64.8 μm) with apophyses and three macroplacoids ( Figs. 27–29 View FIGURES 27 – 29 ). Pharyngeal apophyses distinct, drop-shaped ( Figs. 27–28 View FIGURES 27 – 29 ) (3.7 μm long and 1.2 μm wide in their posterior parts). The first macroplacoid thinner anteriorly, the second oval or slightly rectangular and the third one with a distinct subterminal constriction. The second macroplacoid is distinctly the shortest one (Table 1, Figs. 27–29 View FIGURES 27 – 29 ) and it is evidently placed closer to the first than to the third macroplacoid (in the holotype inter-placoid distances are 2.0 μm and 5.6 μm, respectively). The third macroplacoid is usually slightly longer than the first one (see length ratio macroplacoid 3/macroplacoid 1 in Table 1). The microplacoid is absent, however there is a faint cuticular match-like structure growing out of the posterior end of each of the third macroplacoids. These match-like structures are wider in their terminal parts ( Figs. 27, 29 View FIGURES 27 – 29 ).
Claws of the hufelandi - type, slender ( Figs. 30–31 View FIGURES 30 – 34 , 35–36 View FIGURES 35 – 38 ). Hind claws are slightly longer than claws I–III (Tables 1–2). Primary branches with long, protruding accessory points ( Fig. 30–31 View FIGURES 30 – 34 , 35–37 View FIGURES 35 – 38 ). Lunules on all legs smooth (LM, SEM), better developed on legs IV ( Figs. 32 View FIGURES 30 – 34 , 38 View FIGURES 35 – 38 ). Double transverse cribriform areas below claws of the first three pairs of legs are visible.
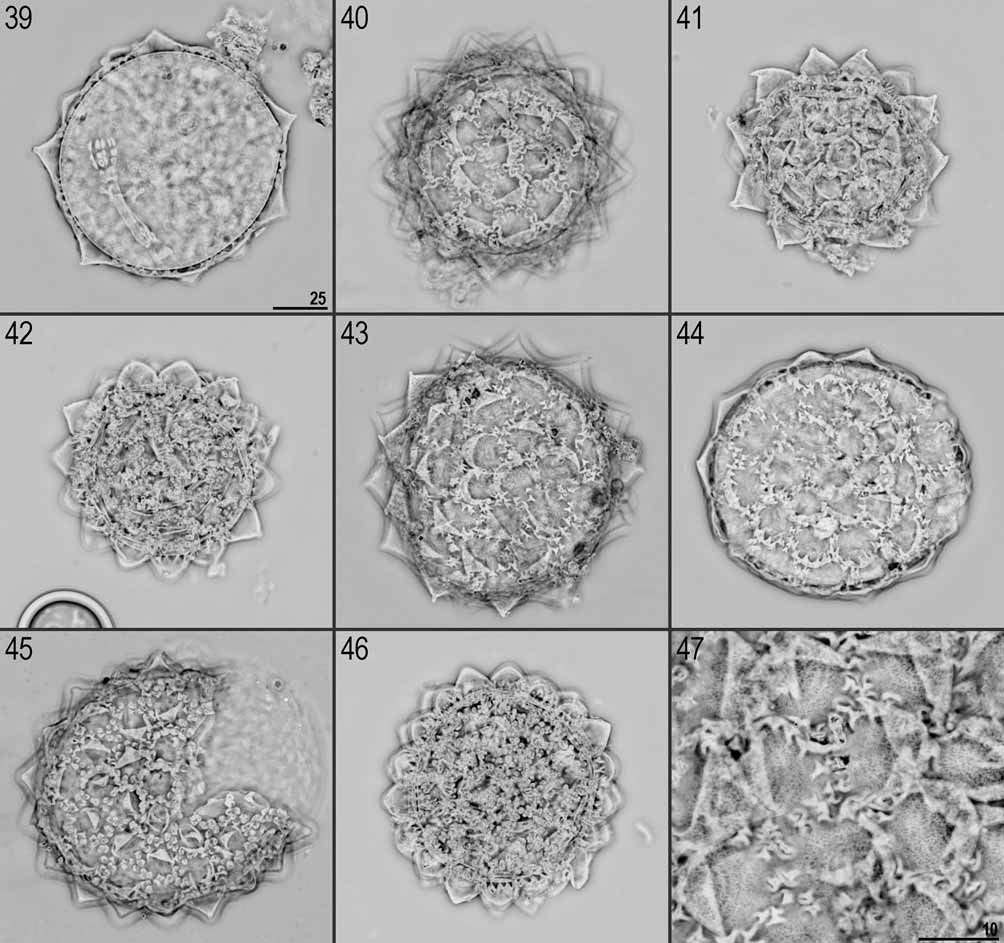
Eggs: White/transparent, laid freely. Spherical or slightly oval, with 12–16 processes on the circumference, areolation around bases of processes present ( Figs. 39–46 View FIGURES 39 – 47 , 48–50 View FIGURES 48 – 51 ).
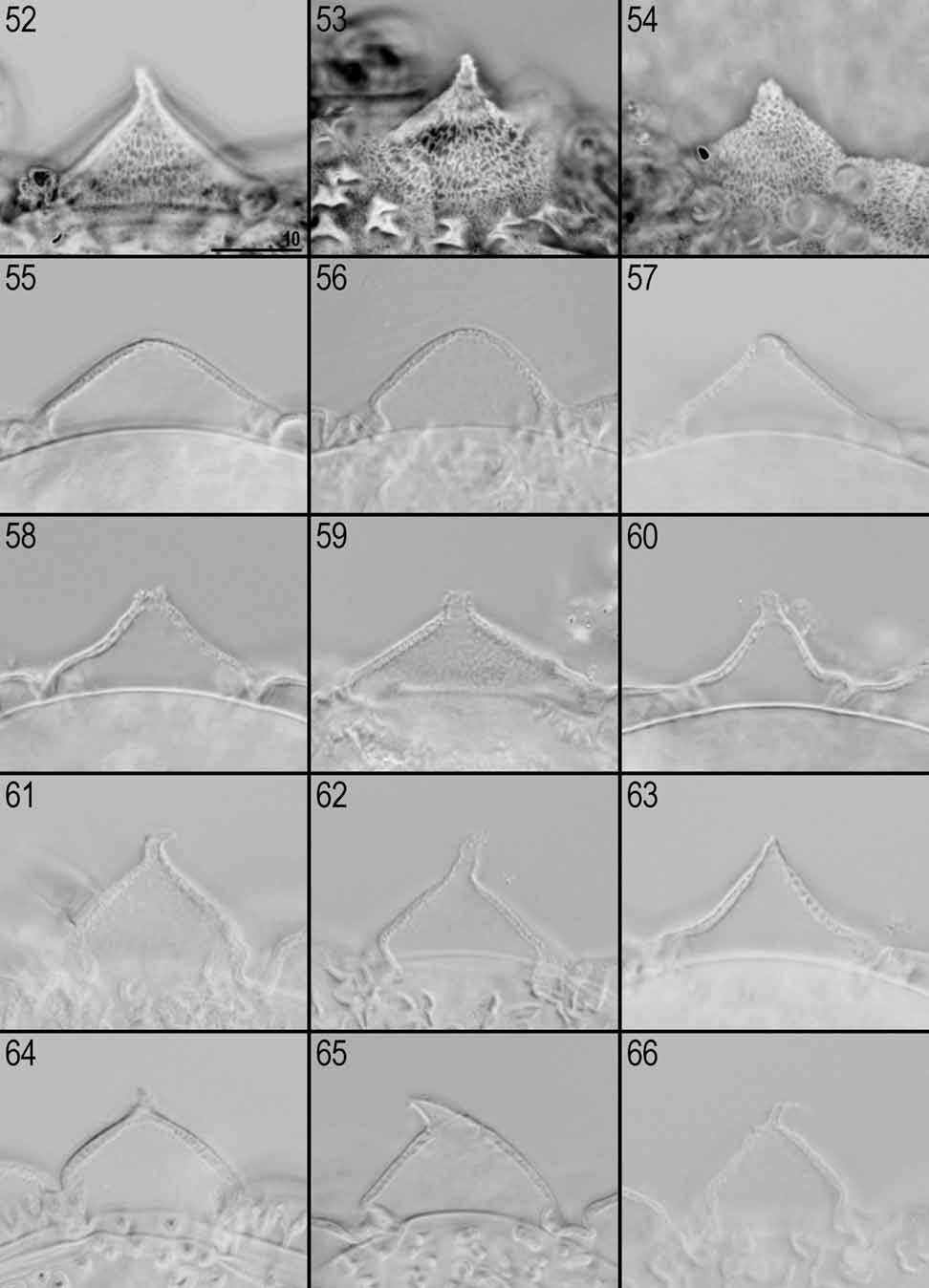
Processes generally in the shape of short and relatively wide cones (the base diameter of each process is larger than the height) with a single short and flexible tip ( Figs. 51–70 View FIGURES 48 – 51 View FIGURES 52 – 66 View FIGURES 67 – 72 ). The tips are variable in their shapes and sizes (sometimes they can be undeveloped or broken). Terminal part of egg processes (including the tip) is usually covered by irregular tubercles poorly visible or invisible in LM ( Fig. 53 View FIGURES 52 – 66 ) but very distinct in SEM ( Fig. 70 View FIGURES 67 – 72 ). Processes consist of double walls with transverse supporting walls forming ‘cells’ visible in PhC and NC as a reticular design ( Figs. 52–54 View FIGURES 52 – 66 ). This reticulation is generally slightly elongated vertically (mesh 0.5–0.9 μm) and can form horizontal rows. External walls of processes without pores ( Figs. 67–69 View FIGURES 67 – 72 ).
The surface between processes is areolated and the ‘reticulation’ of egg processes walls continuously covers the areolae rims. The areolation is of the huziori - type. Mostly oval and narrow areolae are usually transversally divided into two shallow areolae and each of them branches into two smaller and deeper areolae towards the egg surface (some areolae are not branched). The spaces between neighbour areolae are usually broader than the areolae widths. Approximately 12 areolae form a ring around the base of each process ( Figs. 47 View FIGURES 39 – 47 , 51 View FIGURES 48 – 51 , 71–72 View FIGURES 67 – 72 ).












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.