
Japanagromyza parvula Spencer
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3709.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:0EAE3D84-E1E8-442D-A9BD-3E7046A2A706 |
|
DOI |
https://doi.org/10.5281/zenodo.6150014 |
|
persistent identifier |
https://treatment.plazi.org/id/03F68B67-BB1F-E131-71E6-FD61FB13FBAF |
|
treatment provided by |
Plazi |
|
scientific name |
Japanagromyza parvula Spencer |
| status |
|
Japanagromyza parvula Spencer View in CoL
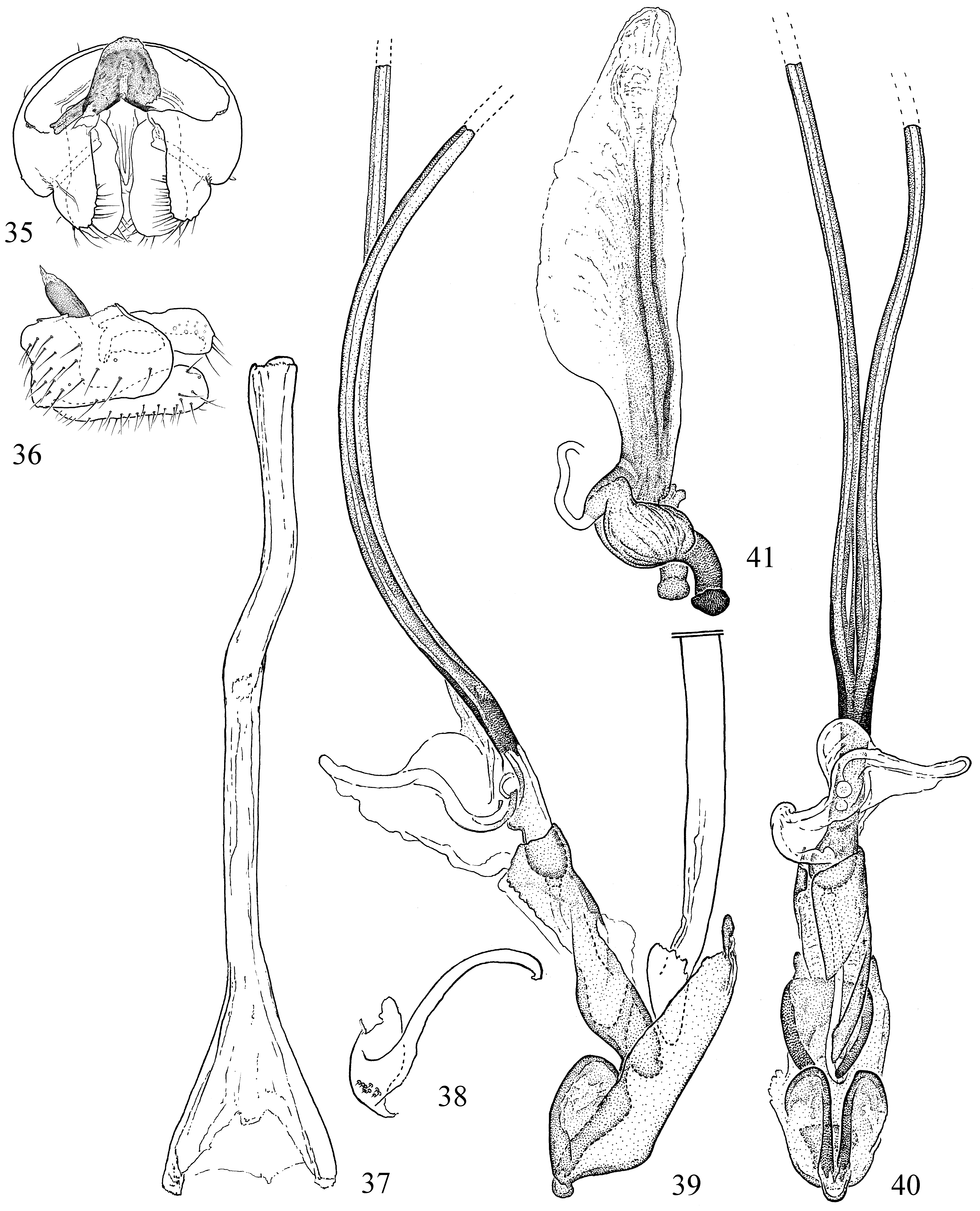
Figs 35–40 View FIGURES 35 – 40
Japanagromyza parvula Spencer 1961a: 1 ; Spencer 1990: 135.
Description. Wing length 2.2–2.6mm (♂), 2.3–2.4mm (♀), 1.9mm (♀ holotype). Length of ultimate section of vein CuA1 divided by penultimate section: 0.6–0.7; vein r-m at 1/3–2/5 length of cell dm. Eye height divided by gena height 5.5–7.6. Lunule with pubescence silvery. Face with slight medial carina.
Chaetotaxy: Third ori sometimes present on one side of frons. Prescutellar acrostichal seta varying from slightly less or more than half length of posterior dorsocentral. Acrostichal setulae usually in 10–12 rows, sometimes 8 (including holotype). Intra post-alar setula-like. Fore tibia with medial seta. Mid femur with several long, curved posterodistal setae. Katepisternum with one smaller additional seta.
Colouration: Setae and setulae dark brown, setulae sometimes paler (including in holotype). Halter white with most of stem brown, and sides of knob with ill-defined brown patches (also see Variation below). Calypter margin and hairs dark brown. Body colour brown with nearly indiscernible green shine.
Genitalia: Subepandrial sclerite bare; narrow arms contiguous with inner margin of surstylus; arms broadly joined medially to form anteriorly projecting plate. Surstylus fused to epandrium, but strongly inset from margin; broadly rounded, half length of epandrium; inner surface setose along midline apically. Cercus large, broad. Hypandrium with extremely elongate apodeme; lateral arms strongly deviated medially at point of contact with postgonite (broken in illustrated male, not drawn). Phallophorus asymmetrical, produced along left margin to point of fusion with basiphallus, which wraps around shaft onto right distal margin. Mesophallus lightly pigmented, tubular, bulging ventroapically, and with base rounded. Hypophallus consisting of membranous folds with curved, projecting tubule that varies from mostly clear to moderately pigmented. Distiphallus with one pair of dark, thin, elongate tubules that join near base. Ejaculatory apodeme with well-sclerotized, bulb-like pileus ejaculatorius that is produced into one pair of curved, heavily sclerotized processes; stem nearly indistinct and basal process reduced; blade narrow, irregular and pale with dark medial rib; base of duct pigmented.
Variation: Some females from RSA and Burundi vary as follows: halter knob entirely white/yellow; orbital plate with glossy spot around base of fronto-orbitals; length of ultimate section of vein CuA1 divided by penultimate section 0.7–0.9; eye height divided by gena height 9.4–15.2; katepisternum with 3 smaller additional setae; posterodistal setae on mid femur less developed; female from Free State with calypter margin and hairs paler brown. One female from RSA (Manguzi Forest Reserve) with entirely white halter excluding brown basal spot on stem and faded brownish spot on knob base. Holotype with halter white excluding brownish stem base.
Host. Fabaceae— Crotalaria agatiflora (Spencer, 1990) .
Distribution. AF: Burundi *, Kenya, Nigeria *, Republic of South Africa *, Tanzania, Uganda *. PA: Oman [?], Saudi Arabia (Deeming, 2006).
Holotype. TANZANIA. “Makoa T.T.O-Afr.”, 9.ii.1959, at light, Lindner (1♀, SMNS).
Material examined. BURUNDI. Burundi Prov.: Réserve Naturelle de Rumonge, 04˚00.940’S, 29˚29.560’E, 17–20.xi. 2012, 900m, A.H. Kirk-Spriggs (2♀, BMSA). NIGERIA. N. Nigeria, Bauchi vegetable garden, 6.iv.1990, J.C. Deeming (1♂, NMWC). RSA. Kwa Zulu-Natal: Kosi Bay Nature Reserve, at(2): 26˚57’17”S, 32˚49’27”E, 13–17.xii.2012, 82m, A.H. Kirk-Spriggs, Malaise traps, woodland savanna (3♂ 3♀, BMSA), Kwa Zulu-Natal: Manguzi Forest Reserve, 26˚59’32”S, 32˚43’25”E, 13–17.xii.2010, 61m, A.H. Kirk-Spriggs (1♀, BMSA), KZN: Ndumo Game R., main camp area at: 26˚54.652’S, 32˚19.719’E, 27–30.xi.2009, A.H. Kirk-Spriggs (4♀, BMSA), Free State: Harrismith Scotland Farm at: 27˚58’59.5”S, 29˚37’09.8”E, 10–12.xi.2009, A.H. Kirk- Spriggs (1♀, BMSA). SAUDI ARABIA. Aseer, Maraba, Malaise trap in fruit farm, 1–17.vi.2003, H.A. Dawah (1♂, NMWC). UGANDA. Entebbe, 7.vi.1972, H. Falke, in forest (1♂, BMNH).
Comments. Japanagromyza parvula and J. tephrosiae are nearly identical in morphology and preliminary findings suggest that their boundaries are in need of redefinition. It is unusual that direct comparisons have never been made in the literature despite their highly derived and nearly identical genitalia, including a very elongate, bifid distiphallus ( Figs 39, 40 View FIGURES 35 – 40 ). The only previously recognized external difference diagnosing the two is the colour of the halter, with that of J. parvula being entirely white and that of J. tephrosiae being brown with the apex becoming white gradually. The present study finds halter pigmentation to be quite variable across African specimens, sometimes even within the same locality—the knob is quite variably brown, often being mostly white on the lateral and sometimes dorsal surfaces (most females), with the margin of the brown sections fading gradually to white (not abruptly delimited, as seen in J. laureata and J. nigrihalterata ).
Aside from this slight variation in pigmentation between Japanagromyza parvula and J. tephrosiae , the only other apparent morphological difference appears to be the relative shape of the external male terminalia ( Figs 35, 36 View FIGURES 35 – 40 ). When viewed laterally, the cercus of an Australian male of J. tephrosiae (figured as J. badia Spencer in Spencer (1977), which was later synonymized by Spencer (1990)) appears to be slightly longer, and the surstylus is longer and considerably narrower; this state is here also assumed to be present in the Indian population as well, as Spencer (1990) noted that the male genitalia of the two sets of reared specimens were “identical”. With these slight differences between the African and Indo /Australian groups, J. parvula is here tentatively redefined to encompass the African populations, and J. tephrosiae is redefined to encompass the Indo /Australian populations.
Host choice within the Fabaceae also varies between the two sets of populations, somewhat supporting this new division. Crotalaria -feeding was recorded for a Kenyan specimen in Spencer (1990) (i.e. Japanagromyza parvula ), and Tephrosia -feeding was recorded in an Australasian population (i.e. J. tephrosiae ). As such, J. tephrosiae is no longer considered to occur in Africa, although additional host records and population sampling are required to support this concept, which is partially based on incomplete published descriptions.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.