
Hydrangea albostellata Samain, Najarro & E.Martínez, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.162.4.1 |
|
persistent identifier |
https://treatment.plazi.org/id/522087A7-FFF1-FFEB-CACE-FF48BDA5FAFC |
|
treatment provided by |
Felipe |
|
scientific name |
Hydrangea albostellata Samain, Najarro & E.Martínez |
| status |
sp. nov. |
Hydrangea albostellata Samain, Najarro & E.Martínez View in CoL , spec. nov., Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4
Mesoamerican functionally dioecious species which has been confused with Hydrangea steyermarkii Standl. , from which it can be distinguished by the following characters: leaves with slightly undulate, spoon-shaped lamina with apiculate apex, abaxially densely white pubescent, especially in young stage, inflorescence axis densely pubescent with adnate, whitish, stellate hairs, calyx lobes of reduced flowers triangular, very small, sometimes nearly invisible, petals membranous, margin revolute.
TYPE: - MEXICO. Chiapas: Pueblo Nuevo Solistahuacan, along the road Pueblo Nuevo Solistahuacan–Rayón , 17°12'41.52"N, 92°57'50.61"W, 1827 m, 19 January 2012, M. S. Samain et al. 2012-023 (holotype: MEXU!, isotypes: CHIP!, GENT!, IEB!, K!, MEXU!, MO!, US!) GoogleMaps [functionally female].
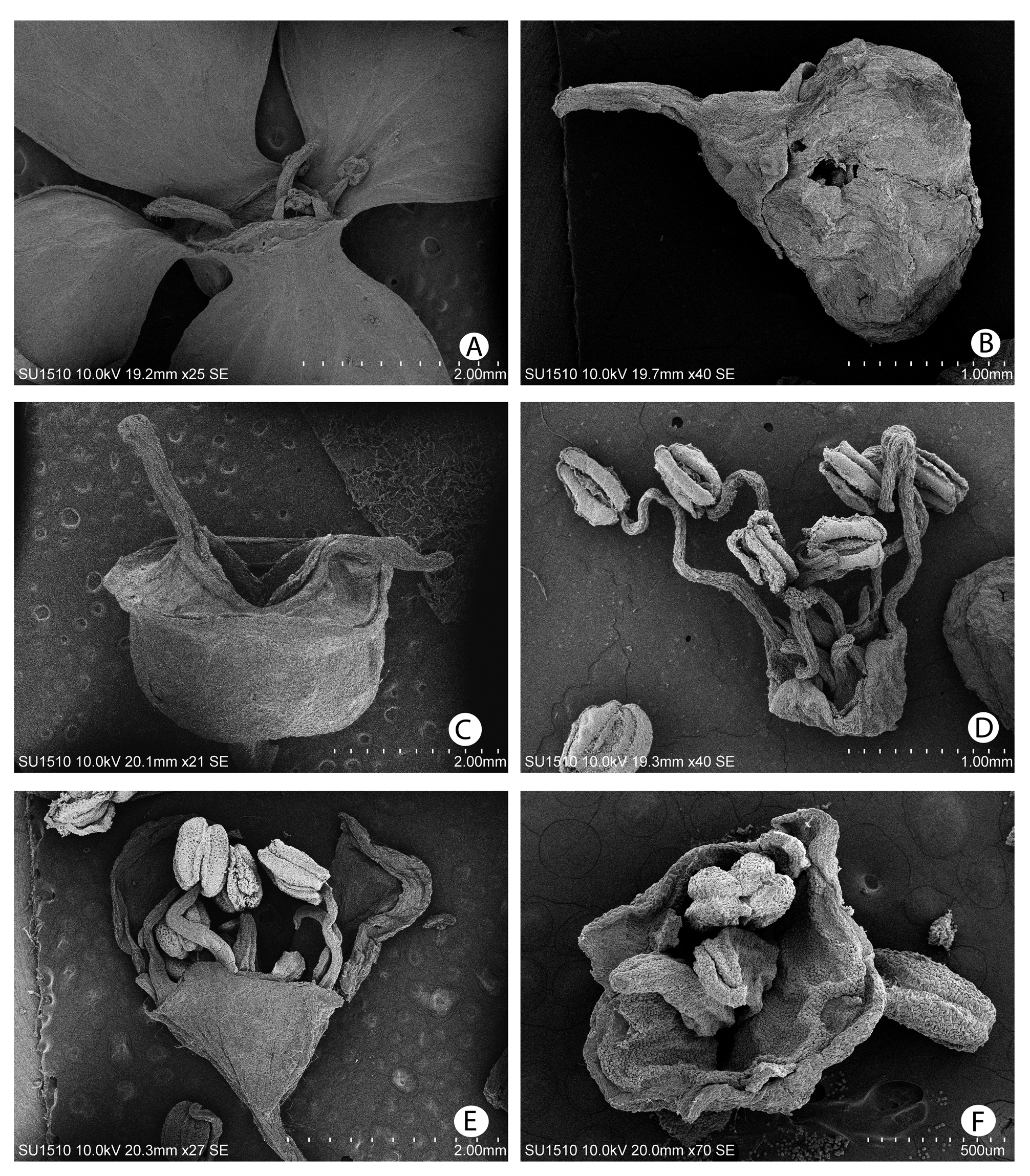
Root-climbing liana of up to 35 m high, funtionally dioecious. Main stem clinging to the host tree with many adventitious roots, up to 20 cm diameter, bark dark brown, with lenticels. Runner shoots (stolons) reddish, with adventitious roots and stellate hairs, with small kataphylls. Branches many-ribbed, slightly angular, with many kataphyll scars, branches attached to host tree with adventitious roots, these roots fibrous, slightly flattened, with caducous stellate indument, free-growing branches terete, slightly fissured and exfoliant. Petiole sulcate adaxially, terete and corchy abaxially, this corchy line sometimes continuing in the abaxial midvein for approximately 1 cm, cortex fragmenting in mature ones, colour brownish grey, when young with whitish penicellate hairs, when mature (nearly) glabrous, 1–3 cm long, leaving a triangular scar on the branch when leaves fall (scar sometimes with slightly rounded borders). Leaves decussate, lanceolate-obovate to elliptic, lamina slightly undulated/spoonshaped, 9–14 cm long, 4–8 cm broad, base cuneate, apex apiculate, leaf margin smooth, venation brochidodromous, veins 6–9, adaxial leaf side with only slightly visible midvein, secondary and tertiary veins very lightly marked, secondary veins join to form submarginal or intramarginal vein, glabrous with few simple, short, white hairs along the basal half of the midvein, abaxially with protruding veins, sparsely pubescent in mature leaves (caducous) and even less hairy on primary and secondary veins, opaque whitish-green, young leaves densely pubescent, slightly reddish, indument caducous except on the midvein, acarodomatia absent, but much indument present in vein axils. Inflorescence axis densely pubescent with adnate, whitish, stellate hairs, 4–15–(25) cm long, robust in functionally female plants, delicate in functionally male plants, with 2–5 opposite or decussate leaf pairs below the inflorescence, the upper two pairs inserted at the base of the inflorescence below the inflorescence bracts, caducous, petiole 2–4 mm long, 3 mm broad, petioles of upper leaf pairs nearly sessile, lamina 0.8–3.5 cm long, 0.7–1.1cm broad, starting from the base of the floral axis, each leaf pair with gradually diminishing size towards the inflorescence, apex of the floral axis woody, cone-shaped, elongated bract scars visible, thickening at the top, 6–8 mm broad, 3–4 mm high in functionally female plants, 5 mm broad, 2 mm high in functionally male plants, inflorescence bracts cucullate, reddish, coriaceous, border membranous, adaxially slightly pubescent, abaxially densely pubescent with whitish stellate hairs, veins nearly invisible because of pubescence, bracts increasing in size, lowermost bract 1.5 cm large, 2 cm broad, higher bracts up to 3.5 cm large, 3 cm broad, consecutively and rapidly caducous during inflorescence development, bracts of partial inflorescences linear, reddish, 8 mm long, 1 mm broad at the base, gradually tapering towards a pointed apex, caducous, inflorescences lateral, opposite, 1–3 pairs of inflorescences per flowering branch, sometimes only 1 inflorescence developing, flowering branch only continues growing vegetatively very slowly during inflorescence development, with maximum two leaf pairs above the inflorescences, or kataphylls only, up to eight pairs, linear, with whitish stellate hairs and a corchy abaxial line, inflorescence axes with basal lignified parts of inflorescences of previous years visible in robust well-collected specimens, allowing to observe growth and flower periodicity, these rests 9–22 cm apart (but relatively constant within one specimen), with 4–9 decussate leaf pairs (or the scars of these leaves, this number constant within one specimen), medulla central in the branch, disappearing in older branches, leaving a hole only, leaves at the base of the inflorescence absent, inflorescence umbellate, buds up to 3.5 cm broad and 3 cm high before opening ( Figs. 1B View FIGURE 1 , 4A View FIGURE 4 ), in flowering stage 5–14 cm diameter, 4–8 cm high, with 3–4 main axes in functionally male plants, 6–9 main axes in functionally female plants, partial inflorescences cymes (when enlarged marginal flowers are present, these terminally placed in each cyme), secondary and tertiary inflorescence axes with whitish stellate hairs, pubescence gradually decreasing towards flower insertion where there are some scarce hairs only. Enlarged marginal flowers sometimes present ( Figs. 3A View FIGURE 3 , 4A, B View FIGURE 4 ), flower pedicel 0.8–1(–1.8) cm long, receptacle rectangular, with 4 lateral ribs, 1.5 mm high, 2.5–3 mm broad, 2 mm diameter, ovary inferior, sepals 4, yellowish to greenish, coriaceous with marked veins, quadrangular with rounded margins to orbicular, sometimes spoon-shaped, margin inwards revolute, (2.5–) 4–8 mm long, (2.5–)4–7(–9) mm broad, petals reduced and caducous, stamens (3–)6, filaments reduced and flattened, 0.3 mm long, 0.2 mm broad, anthers 0.3 mm long, pistils 2, style 1–2 mm long, stigma spathulate to rounded, very shortly penicillate, sometimes with seeds. Flower pedicel of reduced flowers 1.5–2 mm long in functionally male flowers, (2–)3–4(–5) mm long in functionally female flowers, receptacle broadly campanulate in functionally male flowers ( Figs. 3B View FIGURE 3 , 4C View FIGURE 4 ), semiglobose in functionally female flowers ( Figs. 3C View FIGURE 3 , 4D View FIGURE 4 ), ovary inferior, calyx lobes 4, triangular, in some specimens nearly invisible, 0.1–0.3 mm long, seemingly pushing downwards the border of the receptable, petals 4, valvate, cucullate, margin revolute, membranous, 1.8–2.5 mm long, 1.5 mm broad, white; functionally male flowers ( Fig. 3D View FIGURE 3 ): hypanthium 1.5–2 mm diameter to completely round or even quadrangular, stamens up to 9, well-developed, filaments 2.5–4 mm long, anthers 0.7–1 mm long, 0.3–0.5 mm broad, pistils 2, reduced, 0.5–0.6 mm long, stigmas not penicellate; functionally female flowers: stamens 7–9 (if 9: 7 reduced and 2 rudimentary), reduced, filaments 0.1–0.5 mm long, anthers 0.2–0.7 mm long, 0.3 mm broad, pistils 2, 1.2–1.5 mm long, enlarging up to 3 mm during fruit maturation, stigmas apically clavate and shortly penicellate, fruit a semiglobose capsule with 8 lateral ribs ( Figs. 3C View FIGURE 3 , 4D View FIGURE 4 ), apically with a conspicuous border, brownish, (2–)3(– 4) mm high, 3.5 mm broad above, 2–3 mm diameter, opening between the two pistils to release seeds, seeds reddish, elongated, with pointed apices, 0.7–1 mm long, 0.2 mm thick.
Habitat: ―This species is restricted to cloud forest (“selva mediana perennifolia” according to Miranda & Hernández X. (1963), “bosque mesófilo de montaña” according to Rzedowski (1978)) with permanent high humidity, up to 5000 mm rainfall, and high nubosity, on strong to moderate slopes of different geological substrates, metamorphic and sedimentary rocks. It occurs mainly on northeast-facing slopes, where the trade winds strike, in conserved places or on disturbed localities near water streams.
Distribution: ― Hydrangea albostellata is characterized by a wide geographical and altitudinal distribution. In Mexico it occurs in the north and northeast of the state of Chiapas, wherever the required habitat is present, between 1200 and 2300 m above sea level. Interestingly, all recent collections are located between 1800 and 1900 m altitude. Furthermore, this species has been collected in Honduras, El Salvador, Nicaragua and Costa Rica. It is not known from South America.
Phenology: ―In Mexico, this species has been observed flowering from September to January and fruiting in January only. In Central America, plants seems to flower and fruit throughout the year.
Etymology: ―This species can be easily distinguished from all other Mexican and Central American species because of the stellate white pubescence on leaves and inflorescence branches ( Figs. 2B View FIGURE 2 , 4E View FIGURE 4 ). These stellate hairs are partially caducous, which sometimes results in less notoriously pubescent leaves in dried specimens.
Observations: ― Hydrangea albostellata has a larger distribution area than the majority of other species of section Cornidia . This is the only species of southeastern Mexico which had been collected by Dennis Breedlove and his collaborators from the California Academy of Sciences in the framework of the project Flora of Chiapas ( Breedlove, 1986). Nearly all Mexican and Central American collections listed among the additional specimens here studied had been identified on the specimen labels as H. steyermarkii . Three of the five specimens mentioned as H. steyermarkii in the Flora Mesoamericana ( Christenhusz 2009) in fact belong to H. albostellata . See Table 1 for a summary of differentiating characters between both species.
The different duplicates of Breedlove 29829 (see additional specimens examined for details) are functionally female or male, with one specimen (CAS) being mixed. It is highly doubtful that they belong to one and the same individual; leaves of functionally male branches of this collection are narrower and smaller than those of functionally female branches.This is congruent with the 1994 notes on the CAS specimen of Elizabeth McClintock, who worked in the same institute as Breedlove.
As can be seen from the description, the enlarged marginal flowers are not sterile as has been generally assumed, but which we already questioned earlier ( Samain et al. 2010). The description of the marginal flower in this case is from a functionally female plant. The functionally male plants we observed did not have enlarged marginal flowers, showing the intraspecific variability of the presence or absence of this structure in this species, in contrast to e.g. H. seemannii from northern Mexico and H. asterolasia in Costa Rica and northern South America, which always have whitish marginal flowers. Interestingly, we have also observed one enlarged marginal flower in a functionally female plant of H. nebulicola in the Mexican state of Veracruz (M.S. Samain & E. Martinez S. 2011- 215), whereas this was not documented for the functionally male plants in the original description by Nevling & Gómez-Pompa (1968).
The specimen Breedlove 24174 from Solosuchiapa, Chiapas (CAS) belongs to an unidentified species of the family Malpighiaceae , of which juvenile forms have been confused with Hydrangea also in other countries than Mexico, e.g. Salick & Stijfhoorn 8265 (MO!) from Boca de Sábalo, Nicaragua. Nevertheless, even in very young stages, they can be easily distinguished from each other, as leaves of Hydrangea are coriaceous with a dentate to serrate margin, whereas leaves of Malpighiaceae are membranous with a smooth margin. In addition, the specimen from Nicaragua occurs at a very low altitude (± 45 m asl), whereas to our knowledge none of the Hydrangea species in Mesoamerica occurs below 900 m above sea level.
The basal lignified parts of inflorescences and the inflorescence axes of previous years are only visible in robust well-collected specimens, e.g. M.S. Samain & E. Martínez S. 2012-088, 2012-090 ( Fig. 2A View FIGURE 2 ).
Enlarged marginal flowers are sometimes present, e.g. M.S. Samain et al. 2012-021, 2012-023.
Additional specimens examined: ― COSTA RICA. Alajuela: San Ramón, R. B. Monteverde, Cordillera de Tilarán , bosque primario en la Fila Cerros Centinela, 10°17'55"N, 84°47'23"W, 1600 m, 6 June 1994, J. F. Morales 3366 & G. Carnevalli ( INB!) [functionally male] GoogleMaps . Cartago: Cartago, al lado del camino Copatchi–San Rafaël Arriba, en la fila del cerro, 9°49'35.34"N, 84°2'3.93"W, 1890 m, 27 July 2012, M. S. Samain & E. Martínez S. 2012- 033 ( CR!, GENT!, INB!, MEXU!) [dry inflorescences] GoogleMaps ; loc. cit., 1781 m, 9°49'34.81"N, 84°2'9.27"W, 16 January 2013, M. S. Samain & E. Martínez S. 2013-002 ( GENT!, INB!) [dry inflorescences] GoogleMaps . Guanacaste: Tilarán, al lado del camino de Santa Elena al Mirador Lodge , 1.01 km desde la desviación hacia la Reserva Bosque Nuboso Santa Elena, 10°21'10.11"N, 84°48'6.81"W, 1592 m, 31 July 2012, M. S. Samain & E. Martínez S. 2012-049 ( CR!, GENT!, INB!, MEXU!) [inflorescence buds] GoogleMaps ; San Gerardo Abajo, Río Caño Negro, Fincas Quesada y Arce , 10°18'40"N, 84°50'2"W, 1100–1200 m, 05 December 1991, E. Bello 4286 & E. Cruz ( CR!, INB!, MO!) [functionally female, fruits] GoogleMaps . Puntarenas: Puntarenas, Ojo de Agua, Monteverde, Finca Méndez , Río Aranjuez , 10°17', 84°46', 1550 m, 14 November 1987, W. Haber 7728 & E. Bello ( CR!) [functionally female, fruits] ; Monteverde, Pacific slope above Quebrada Máquina , 10°18'N, 84°48'W, 1400 m, 26 September 1991, W. Haber 10885 ( CR!, INB!, MO!) [flower buds] GoogleMaps ; Monteverde, Quebrada Máquina upstream from Hotel Belmar , 10°18'N, 84°48'W, 1400 m, 17 December 1991, W. Haber 10935 & W. Zuchowski ( CR!, MO!) [functionally female, fruits] GoogleMaps ; upper San Luis River Valley on Pacific slope below Monteverde, 10°20'N, 84°50'W, 1300–1400 m, 26 August 1986, W. A. Haber ex E. Bello C., A. Clagget & M. Powell 5398 ( INB!) [functionally male] GoogleMaps ; San Luis, Monteverde, camino a Veracruz , 10°16'35"N, 84°47'45"W, 1040–1060 m, 16 October 1992, A. Fernández 438 ( INB!) [flower buds] GoogleMaps ; R. B. Monteverde, Cordillera de Tilarán, San Luis, Finca Juan Ramón Fuentes, Buen Amigo , 10°16'33"N, 84°47'45"W, 1220 m, 10 December 1993, Z. Fuentes 445 & E. Fuentes ( CR!, INB!) [functionally male] GoogleMaps ; R. B. Monteverde, Cordillera de Tilarán, San Luis de Monteverde, bosque en la cuenca de San Luis, 10°16'33"N, 84°47'45"W, 1100 m, 15 August 1993, K. Taylor 112 ( CR!, INB!, MO!) [inflorescence buds] GoogleMaps ; Monteverde Reserve, high peaks near TV towers, 10°20'N, 84°50'W, 1775 m, 11 October 1985, W. A. Haber ex E. Bello 3070 ( CR!, MO!; label on MO specimen only mentions E. Bello 3070) [functionally male] GoogleMaps . San José, Escazú, ZP Cerros de Escazú, Cuenca del Tárcoles , San Antonio , Bebedero , Cercanías de las antenas, La Ventolera , a orilla del camino, 9°53'27"N, 84°9'22"W, 1784–1800 m, 10 September 2005, D. Santamaria 3001 & R. Aguilar ( INB!) [inflorescence buds] GoogleMaps . HONDURAS. Lempira: alrededor de el Río Naranjo y bajo el Campamento Don Tómas. Parque Nacional de Celaque, 14°33'N, 88°40'W, 2000 m, 27 May 1991, P. House 996 ( CAS!, EAP, HEH, MO!, TEFH) [functionally female, fruits] GoogleMaps ; loc. cit., 13 November 1991, P. House 1105 ( EAP, HEH, MEXU!, MO (2)!, TEFH) [functionally male] GoogleMaps . Yoro: Río Pijol Valley, 7 km south east of Nueva Esperanza, along forested Quebrada that flows into Río Pijol , adjacent slopes, and coffee finca above, 15°12'N, 87°35'W, 1300–1500 m, 30 May 1993, R. L. Liesner 26699 ( MO) [functionally female, fruits] GoogleMaps . EL SALVADOR. Volcán de San Salvador, 1000–1800 m, 7 April 1922, P. C. Standley 22966 (US(2)!) [juvenile] . MEXICO. Chiapas: La Trinitaria, 4 km east of Laguna Tsiskaw [Tziscao] near Dos Lagos , 1300 m, 19 October 1974, D. E. Breedlove 38792 ( CAS!, DUKE!, MO!, NY!) [functionally male] ; loc. cit., between Lago Tziscao and Dos Lagos, Lagos de Montebello National Park , 1372 m, 13 October 1981, D. E. Breedlove 53460 ( CAS!, MO!) [functionally male] ; slopes east of Laguna Tzikaw [Tziscao], Monte Bello National Park, 1300 m, 23 January 1973, D. E. Breedlove 32250 & A. R. Smith ( CAS!) [functionally female, mature fruits] ; Rayón, steep slope in the Selva Negra 10 km above Rayón Mezcalapa along road to Jitotol , 1700 m, 25 November 1972, D. E. Breedlove 29829 & R. L. Dressler ( CAS! Mixed functionally male and female), DUKE! [294758- ♂, 294765- ♀], MEXU! ♀ (immature fruits) , MICH! ♂ , MO! [3884984- ♂, 3884985- ♀] , US! ♀ (flowers (buds) + immature fruits) [different duplicates are functionally female or male]; loc. cit., 9 miles northwest of Pueblo Nuevo Solistahuacan along road between Rincon Chamula and Rayón , 5800 feet, September 1971, R. F. Thorne & E. Lathrop 41591- A ( CAS!) [inflorescence buds] ; Tenejapa, Paraje Shohleh , 8400 feet, 12 January 1966, A. Shilom Ton 573 ( MO!) [functionally female, immature fruits] ; La Independencia, third ridge along road from Las Margaritas to Campo Alegre , 2300 m, 5 November 1984, D. E. Breedlove 62118 ( CAS!) [functionally male] ; loc. cit., 5 November 1984, D. E. Breedlove 62117 ( CAS!, MICH!) [functionally female, flowers + immature fruits] ; Jitotol, about 12 km north of Jitotol along a side road to an oil well, 2000 m, 28 September 1971, D. E. Breedlove 19944 ( CAS!) [juvenile] ; Pueblo Nuevo Solistahuacan, along the road Pueblo Nuevo Solistahuacan–Rayón, 17°12'40.88"N, 92°57'49.71"W, 1848 m, 19 January 2012, M. S. Samain et al. 2012- 022 ( CHIP!, GENT!, IEB!, MEXU!) [functionally female, young fruits] GoogleMaps ; loc. cit., 17°12'40.55"N, 92°57'48.53"W, 1850 m, 19 January 2012, M. S. Samain et al. 2012-024 ( CHIP!, GENT!, MEXU!) [fallen bracts only] GoogleMaps ; Arroyo Grande–Pueblo Nuevo Solistahuacan, 17°10'53.64"N, 92°52'34.94"W, 1878 m, 19 January 2012, M. S. Samain et al. 2012-021 ( CHIP!, GENT!, IEB!, MEXU!) [functionally female, immature fruits] GoogleMaps ; loc. cit., 9 September 2012, M. S. Samain & E. Martínez S. 2012-089 ( CHIP!, GENT!, IEB!, MEXU!) [functionally female] GoogleMaps ; Tapalapa, camino Coapilla–Ocotepec, near Río Quachi , 17°11'35.91"N, 93°7'51.81"W, 1862 m, 3 January 2012, M. S. Samain et al. 2012-005 ( CHIP!, CR!, GENT!, GUAT!, IEB!, MEXU!, MO!, UAMIZ!, US!) [functionally male] GoogleMaps ; loc. cit., 9 September 2012, M. S. Samain & E. Martínez S. 2012-090 ( CHIP!, GENT!, IEB!, MEXU!) [functionally male] GoogleMaps ; loc. cit., 17°11' 36.37"N, 93°7'56.74"W, 1736 m, 3 January 2012, M. S. Samain et al. 2012-006 ( CHIP!, GENT!, IEB!, MEXU!, QMEX!) [functionally female, mature fruits] GoogleMaps ; Yajalón, La Ventana, 17°9'59.17"N, 92°23'50.90"W, 1843 m, 6 September 2012, M. S. Samain et al. 2012-085 ( CHIP!, GENT!, MEXU!) [sterile depauperated individual] GoogleMaps ; loc. cit., 17°9'24.36"N, 92°22'42.01"W, 1823 m, 6 September 2012, M. S. Samain et al. 2012-088 ( CHIP!, GENT!, MEXU!) [functionally male] GoogleMaps ; Tapalapa, Cerro de la Bandera, 17°12'37.41"N, 93°5'49.94"W, 1937 m, 19 August 2011, M. S. Samain et al. 2011-201 ( CHIP!, GENT!, MEXU!) (juvenile) GoogleMaps . NICARAGUA: Matagalpa, Cordillera Central of Nicaragua between Aranjuez and Peor es Nada , 1500 m, 1 November 1968, A. Molina R. 22965 ( MO!) [functionally male] ; Mountain El Arenal, road to Aranjuez , 1300 m, 7 January 1977, A. Molina R. 31589 ( MO!) [functionally male] . Jinotega, road between Jinotega and Matagalpa, La Galia, 1600 m, 6 December 1958, J. G. Hawkes , J. P. Hjerting & R. N. Lester 2183 ( K!) [juvenile] .
Conservation: ―According to the criteria and categories of IUCN (2012), H. albostellata should be considered as endangered, but the current intrinsic threats of its habitat will soon drive it to be critically endangered, especially as a consequence of habitat destruction. The population in the municipality of La Trinitaria is probably extinct as a consequence of fires during the El Niño in 1997 and 1998. It had not rained since October 1997 for 8 months, when in May 1998 fires allover the state of Chiapas destroyed 250,000 hectares of primary forest ( Martínez, 2003). The annual precipitation in La Trinitaria then averaged 5000 mm, with daily rains, so it is even quite probable that all plants died before the fires, as was observed in the region of the Chimalapas in eastern Chiapas and in western Oaxaca (Silvia Salas, pers. comm.). The population in the municipality of La Independencia could not yet be found, but it is highly probably that some individuals still survive in the remaining patches of primary cloud forest. The population in the municipality of Pueblo Nuevo Solistahuacan has been detected and seems viable, despite the fact that many mature individuals grow in primary forest areas near the roads. During our exploring expeditions, new populations have been discovered in the municipalities of Tapalapa and Yajalón, both in local reserves, guaranteeing their survival for at least some time. Taking into consideration its current extensive but fragmented distribution area, it was probably once a relatively common species in humid cloud forest from southeastern Mexico to Costa Rica.



| M |
Botanische Staatssammlung München |
| S |
Department of Botany, Swedish Museum of Natural History |
| MEXU |
Universidad Nacional Autónoma de México |
| CHIP |
Instituto de Historia Natural |
| GENT |
Ghent University, Biology Department |
| IEB |
Instituto de Ecología, A.C. |
| K |
Royal Botanic Gardens |
| MO |
Missouri Botanical Garden |
| R |
Departamento de Geologia, Universidad de Chile |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| J |
University of the Witwatersrand |
| F |
Field Museum of Natural History, Botany Department |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| INB |
Instituto Nacional de Biodiversidad |
| E |
Royal Botanic Garden Edinburgh |
| CR |
Museo Nacional de Costa Rica |
| W |
Naturhistorisches Museum Wien |
| A |
Harvard University - Arnold Arboretum |
| C |
University of Copenhagen |
| Z |
Universität Zürich |
| TV |
Centro de Estratigrafia e Paleobiologia da Universidade Nova de Lisboa |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| CAS |
California Academy of Sciences |
| EAP |
Escuela Agrícola Panamericana |
| HEH |
Escuela Nacional de Ciencias Forestales |
| TEFH |
Universidad Nacional Autónoma de Honduras |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| DUKE |
Duke University |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| MICH |
University of Michigan |
| GUAT |
Museo Nacional de Historia Natural |
| UAMIZ |
Universidad Autónoma Metropolitana, Iztapalapa |
| QMEX |
Universidad Autónoma de Querétaro, Centro Universitario |
| N |
Nanjing University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |