
Hyalinobatrachium mashpi, Guayasamin & Brunner & Valencia-Aguilar & Franco-Mena & Ringler & Medina Armijos & Morochz & Bustamante & Maynard & Culebras, 2022
|
publication ID |
https://doi.org/ 10.7717/peerj.13109 |
|
publication LSID |
lsid:zoobank.org:pub:0C4888D5-2DB9-4421-A96E-7E41C17EC82F |
|
DOI |
https://doi.org/10.5281/zenodo.6377362 |
|
persistent identifier |
https://treatment.plazi.org/id/0815B7E6-33FB-42D9-A367-4FB50885C256 |
|
taxon LSID |
lsid:zoobank.org:act:0815B7E6-33FB-42D9-A367-4FB50885C256 |
|
treatment provided by |
Valdenar |
|
scientific name |
Hyalinobatrachium mashpi |
| status |
sp. nov. |
Hyalinobatrachium mashpi new species
LSID: 0815B7E6-33FB-42D9-A367-4FB50885C256
Suggested English name: Mashpi Glassfrog
Suggested Spanish name: Rana de Cristal de Mashpi
Holotype. CJ11642 , adult male from San Vicente River (0.16334 N, 78.86736 W; 1,040 m a.s.l.), Mashpi Reserve , Pichincha Province, Ecuador, collected by Jaime Culebras and Carlos Morochz on 28 September 2019. GoogleMaps
Paratopotypes . CJ11643–44 , adult males with same data as holotype GoogleMaps .
Paratypes. MZUTI-3921 , adult male from Amagusa River (0.15469 N, 78.85322 W; 1,137 m a.s.l.), Amagusa Reserve , Pichincha Province, Ecuador, collected by Carlos Morochz and Lucas Bustamante on 14 December 2014. CJ11645, GoogleMaps adult male from tributary of the Mashpi River (0.11463 N, 78.88307 W; 1,126 m a.s.l.), Tayra Reserve , Pichincha Province, Ecuador, collected by Jaime Culebras on 28 October 2019. GoogleMaps
Generic placement. The new species is placed in the genus Hyalinobatrachium Ruiz-Carranza & Lynch (1991) , as modified by Guayasamin et al. (2009), on the basis of morphological and molecular data. The molecular phylogeny ( Fig. 2 View Figure 2 ) places the new species within the genus Hyalinobatrachium with high confidence. Phenotypically, the main diagnostic traits of Hyalinobatrachium are: (1) completely transparent ventral parietal peritoneum; (2) digestive tract and bulbous liver are covered by iridophores; (3) absent humeral spines; (4) white bones in life; (5) males call from the undersides of leaves; (6) females place the eggs on the undersides of leaves; (7) males provide extended parental care; and (8) tympanum with an orientation that places it almost on a horizontal plane (instead of a more lateral plane as observed in other glassfrog genera). All the aforementioned characteristics are present in Hyalinobatrachium mashpi sp. nov. We note that we have observed males on the same leaves as egg clutches for continuous days, but additional studies are necessary to confirm that these observations actually represent extended paternal care.
Diagnosis. Hyalinobatrachium mashpi sp. nov. is distinguished from other species in the genus mainly by its dorsal coloration (i.e., head with light yellow spots that may form an interorbital bar; dorsum lime green with small light yellow spots) and by its transparent pericardium (i.e., red heart visible in ventral view). Hyalinobatrachium mashpi sp. nov. is most similar to H. aureoguttatum , H. talamancae , H. valerioi , H. vireovittatum , and
the new species described below. Differences among these species are indicated in Table 1 View Table 1 and Figs. 3–5 View Figure 3 View Figure 4 View Figure 5 . The new species is morphologically cryptic with Hyalinobatrachium nouns sp. nov. (described below); however, the two new species exhibit a considerable genetic distance (16S; 4.6–4.7%), which is particularly remarkable given that they are found at relatively close geographic proximity (straight distance = 18.9 km), but separated by the Intag-Guayllabamba river valley.
Characterization. The following combination of characters are found in
Hyalinobatrachium mashpi sp. nov.: (1) dentigerous process of the vomer lacking teeth; (2) snout truncate in dorsal view and slightly protruding in lateral view; (3) tympanum oriented almost horizontally; tympanic annulus barely visible, hidden under skin; tympanic membrane differentiated, with coloration similar to that of surrounding skin; (4) dorsal skin shagreen; (5) ventral skin areolate; cloacal ornamentation absent, paired round tubercles below vent absent; (6) parietal peritoneum and pericardium translucent (in life, red heart visible in ventral view); liver, viscera and testes covered by iridophores; (7) liver bulbous; (8) humeral spines absent; (9) hand webbing formula: I (2 + –2 1/2)—(3 – –3) II (2–2 –)—(3 + –3 1/4) III (2–2 +)—(1 3/4 –2) IV; (10) foot webbing moderate; webbing formula: I 1 + —(2–2 +) II (1–1 1/3)—2 1/4 III (1 1/3 –1 1/2) —(2 + –2 1/4) IV (2 1/2 –2 1/3)—1 V; (11) fingers and toes with thin lateral fringes; ulnar and tarsal folds absent; (12) nuptial excrescence present as a small pad on Finger I (Type V), prepollex not enlarged; prepollical spine not projecting (spine not exposed); (13) when appressed, Finger I longer than II;
(Continued)
(14) diameter of eye about 2 times wider than disc on Finger III; (15) coloration in life: dorsal surfaces lime green with small light yellow spots; (16) coloration in preservative: dorsal surfaces creamish white, with minute lavender melanophores; (17) eye coloration in life: iris yellow to golden-yellow; pupil surrounded by lavender ring; (18) melanophores absent from fingers and toes, except Toes IV and V; (19) males call from underside of leaves; advertisement call consisting of single pulsed note, with duration of 0.37–0.46 s, peak frequency at 5.25–5.60 kHz, maximum frequency at 5.46–5.81 kHz, and minimum
frequency at 4.62–4.92 kHz; (20) males attend egg clutches located on the underside of leaves overhanging streams; clutch size of 31 or 32 embryos (n = 2); (21) SVL in adult males 19.7–20.9 mm (mean = 20.5; n = 5); females unknown; and (22) enameled tubercles absent from sides of head.
Description of the holotype. CJ11642, adult male with SVL 20.6 mm. Head wider than long (head width 39% of SVL; head length 78% of head width). Snout truncate in dorsal view and slightly protruding in lateral view. Loreal region concave, nostrils slightly protuberant, elliptical; internarial region concave; canthus rostralis not well defined. Eyes
small, directed anterolaterally, eyes about 50 Ǫ relative to midline (where anterior-facing eyes would be 90 Ǫ relative to midline). Tympanum barely visible, oriented almost horizontally; tympanic membrane differentiated and pigmented as surrounding skin. Dentigerous processes on vomers absent; choanae large, oval, separated widely (distance about the same as between nostrils); tongue round, white in preservative, anterior 4/5 attached to mouth; posterior border of tongue widely notched; vocal slits present, extending along floor of mouth lateral to tongue; enameled glands absent from lower part of upper jaw. Ulnar fold absent; humeral spine absent. Relative length of fingers:
II <I <IV <III; finger discs rounded, about the same size as discs on toes, disc on Finger III 42% of eye width; hand webbing reduced between Fingers I–III, moderate between Fingers III and IV, with formula I 2 + —3 – II 2 – —3 1/5 III 2 + —1 3/4 IV. Prepollex concealed; subarticular tubercles round, faint; few small supernumerary tubercles present, palmar tubercle round, of moderate size and difficult to see, thenar tubercle ovoid; nuptial excrescences present as a small pad on external edge of Finger I (Type V). Hind limbs slender, tibia length 55% of SVL; tarsal fold absent; discs of toes round; inner metatarsal tubercle small, outer metatarsal tubercle round, both very difficult to distinguish. Webbing formula of feet: I 1 + —2 II 1—2 1/4 III 1 1/2 —2 + IV 2 1/2 —1 V. In preservative, dorsal skin creamish white, with minute dark lavender melanophores (only visible under the stereomicroscope); dorsal skin shagreen; skin on venter areolate; cloacal opening at level of upper thighs, small and non-enameled cloacal warts present. Parietal peritoneum and pericardium translucent (in life, the red heart is visible ventrally); urinary bladder lacking iridophores; liver, viscera, and tested fully covered by iridophores. Kidneys rounded, approximately bean-shaped; liver bulbous.
Coloration in life. Dorsal surfaces apple green to yellowish green with diffuse yellow spots; head with light yellow spots that may form an interorbital bar. Melanophores absent from fingers and toes, except Toes IV and V. Ventrally, parietal peritoneum and pericardium transparent, with a red heart always visible. Gall bladder and urinary bladder covered by translucent peritonea; hepatic and visceral peritonea covered by white iridophores; ventral vein red. Iris yellow, with numerous minute lavender spots. Bones white.
Coloration in preservative. Dorsal surfaces creamish white dotted with minute dark lavender melanophores; venter uniform cream, with partial translucence; pericardium translucent; visceral peritoneum covered by iridophores. Iris white with minute lavender melanophores. Melanophores absent from hands and feet, except from some few present on dorsal surfaces of Toes IV and V.
Measurements of holotype (in mm). CJ11642, adult male. SVL = 20.6, femur
length = 11.4, tibia length = 11.3, foot length = 9.6, head length = 6.2, head width = 8.0,
interorbital distance = 2.4,eye diameter = 2.6, tympanum diameter = 0.6, arm length = 4.2, hand length = 6.3, Finger I length = 4.6, Finger II length = 4.1, width of Finger III = 1.1.
Vocalizations ( Figs. 6 View Figure 6 , 7 View Figure 7 ). We measured call variables from two individuals, each from a different locality, Mashpi Reserve (CJ11642; call code LBE-C-051) and Tayra Reserve (CJ11645; call code LBE-C-052). The call of Hyalinobatrachium mashpi sp. nov. ( Fig. 6 View Figure 6 )
consists of a single pulsed (amplitude-modulated) note, which starts with one lower-frequency pulse followed by ~9 more consistent pulses at a slightly higher dominant frequency. We analyzed variables from both individuals: four calls from CJ11645 and eight calls from CJ11642. Calls in our field recordings had a duration of 0.373–0.461 s
(mean = 0.425 ± 0.027 SD, n = 12). Time between calls ranged from 10.07–17.48 s (mean = 12.80 ± 2.166 SD, n = 10); intervals between H. mashpi calls were longer when a sympatric glassfrog ( Espadarana prosoblepon ) called in the interim period. Peak frequency was 5.25–5.6 kHz (mean = 5.38 kHz ± 0.12 SD; n = 12), with a maximum frequency of 5.46–5.81 kHz (mean = 5.38 kHz ± 0.11 SD; n = 12) and a minimum frequency of 4.62–4.92 kHz (mean = 4.79 kHz ± 0.10 SD; n = 12).
We compared the calls of H. mashpi sp. nov. to those of phenotypically and/or genotypically similar species within the same genus: H. adespinosai , H. aureoguttatum , H. pellucidum , H. tatayoi , and H. valerioi ( Table 2 View Table 2 ). The call of H. adespinosai is a single pulsed (amplitude-modulated) note, consisting of ~12 pulses (mean call duration = 0.54 s ± 0.007 SD, n = 10). Time between calls ranged from 10.87–30.04 s (mean inter-call interval = 20.12 s ± 8.77 SD). Mean peak/fundamental frequency was 4.94 kHz (±0.07 SD; range = 4.87–5.04 kHz), with a mean maximum frequency of 5.11 (±0.08 SD; range = 5.0–5.25 kHz) and a mean minimum frequency of 4.57 (±0.15 SD; range = 4.32–4.75 kHz). The call of H. aureoguttatum ( Fig. 6 View Figure 6 ; Table 2 View Table 2 ) consists of a very short, single tonal note (mean call duration = 0.082 s ± 0.002 SD, n = 24). Time between calls ranged from 1.99–5.20 s (mean inter-call interval = 3.24 s ± 0.79 SD, n = 23). Mean peak/fundamental frequency was 6.86 kHz (±0.25 SD; range = 6.55–7.41 kHz; n = 24 calls). Two harmonics are present. We measured call variables from individuals recorded in Canandé (0.5112 N, 79.1343 W; 457 m), Esmeraldas Province, Ecuador, in December 2018 by AVA (LBE- 053–55), and in Mashpi Lodge Reserve (0.17057 N, 78.888 W; 721–723 m) in March 2019 by RMB (LBE-056, 057).
The call of H. chirripoi is a single pulsed (amplitude-modulated) note, consisting of ~12 pulses (mean call duration = 0.255 s ± 0.03 SD, n = 2). Since our recording only included two bouts of calling, we were unable to include H. chirripoi in the DAPC analysis.
The interval between the two calls was 84.3 s. Peak/fundamental frequency was 4.48 kHz, with a maximum frequency of 4.99–5.77 kHz and a minimum frequency of 4.16–4.21 kHz. We measured call variables from one individual recorded in Reserva Itapoa (0.51307 N, 79.134 W; 321 m), Esmeraldas Province, Ecuador, in July 2016 by JC (LBE-019).
The call of H. pellucidum consists of a short, single tonal note (mean call
duration = 0.129 s ± 0.009 SD, n = 41). Time between calls ranged from 1.67–5.35 s (mean inter-call interval = 2.94 s ± 0.79 SD). Mean peak/fundamental frequency was 5.70 kHz (±0.06 SD; range = 5.60–5.86 kHz), with a mean maximum frequency of 6.0 (± 0.06 SD, range 5.86–6.14 kHz) and a mean minimum frequency of 5.16 (±0.07 SD, range 5.05–5.32 kHz). We measured call variables from one individual (USNM 286708) recorded at Río Azuela, Napo Province, Ecuador, by Roy McDiarmid on February 23th, 1979.
The call of H. tatayoi consists of a short, single tonal note (mean call duration = 0.143 s ± 0.04 SD, n = 26). Time between calls ranged from 2.05–21.68 s (mean inter-call interval = 7.64 s ± 4.92 SD). Mean peak/fundamental frequency was 4.82 kHz (±1.77 SD; range = 4.45–5.11 kHz), with a mean maximum frequency of 5.14 (±0.17 SD, range 4.83–5.40 kHz) and a mean minimum frequency of 4.24 (±0.34 SD, range 3.30–4.61 kHz). We measured call variables from four individuals recorded in Jama Coaque Reserve (0.108264 S, 80.117701 W; 700 m), Manabí Province, Ecuador, in March 2019 by RMB.
The call of H. valerioi ( Fig. 6 View Figure 6 ) consists of a single tonal note (mean call duration = 0.079 s ± 0.01 SD, n = 70). Time between calls ranged from 1.76–8.00 s (mean inter-call interval = 4.27 s ± 1.2 SD). Mean peak frequency was 6.77 kHz (±0.19 SD;
range = 6.46–7.24 kHz). Harmonics are likely present but are difficult to discern in the available recordings. We measured call variables from three individuals recorded in Costa Rica (Limón and Rincón de Oso) by Roy McDiarmid. We used the following recordings from the Macaulay Library at the Cornell Lab of Ornithology: ML212787, ML212788, and ML213430.
Results from the discriminant analysis of principal components (DAPC) revealed that the calls of H. mashpi sp. nov. cluster separately, and are thus acoustically distinct from H. adespinosai , H. aureoguttatum , H. pellucidum , H. tatayoi , and H. valerioi ( Fig. 7 View Figure 7 ).
Overlap occurred between H. aureoguttatum and H. valerioi clusters, as well as between H. tatayoi and H. adespinosai clusters. This suggests that the calls of these pairs may not be adequate for species identification alone; more field recordings with genetic verification of
the calling species are thus recommended for future studies. Nearly all (99.9%) of the variance was retained by three principal components. Table 3 View Table 3 lists the eigenvalues and variable loadings of each principal component.
Natural history. Most individuals of Hyalinobatrachium mashpi sp. nov. were found on the underside of leaves among riverine vegetation ( Figs. 8 View Figure 8 , 9 View Figure 9 ). These frogs are difficult to observe because they are found 3–14 m above ground along steep creeks. Males have been observed calling in the months of April, May, June, August, September, October and November. Males that were guarding egg clutches while calling were observed during the rainy season (18 February 2019; 7 May 2021) and dry season (October 2014, June 2015, and August 2021). Examined egg clutches contain 31–34 eggs (n = 3). A female with mature eggs visible through the skin was observed on 27 May 27 2015.
Distribution ( Fig. 10 View Figure 10 ). Hyalinobatrachium mashpi sp. nov. is only known from the following localities in the Mashpi river basin, Pichincha Province, Ecuador: (i) Mashpi Lodge Reserve (San Vicente River, 1,040–1,101 m; Laguna River, 1,069 m); (ii) Amagusa Reserve (Amagusa River, 1,137 m; Mashpi Chico River , 1,130 m); and (iii) Tayra Reserve, 976–1,126 m. Unidentified and closely related frogs from Colombia (Departamento de Risaralda, MAR 2147; Departamento de Valle del Cauca; MAR 2222; Fig. 2 View Figure 2 ) may prove to be conspecifics of H. mashpi .
Evolutionary relationships. Our phylogenetic analyses ( Fig. 2 View Figure 2 ) reveal Hyalinobatrachium mashpi sp. nov. as sister to undetermined haplotypes from the Colombian Andes
(MAR 2147, 2222) and a new species from the Toisán Mountain Range, described below. Other closely related taxa are endemic to Central America: H. vireovittatum and H. talamancae ( Fig. 2 View Figure 2 ).
Etymology. The specific epithet mashpi is used as a noun in apposition and refers to the Mashpi area in northwestern Ecuador. There are several conservation efforts to preserve the last patches of forest remaining in Mashpi (e.g., Mashpi Lodge Reserve, Tayra Reserve, Amagusa Reserve, Mancomunidad del Chocó Andino, Chocó Andino Biosphere Reserve). Mashpi is a Yumbo word that means ‘friend of water’, an apt description of this glassfrog, which depends on healthy streams for its reproduction.
Conservation status. We recommend that Hyalinobatrachium mashpi be listed as Endangered, following IUCN Red List criteria B1ab(iii): extent of occurrence estimated to be less than 5,000 km 2; known to exist at no more than 5 localities; and continuing decline, observed, inferred or projected, in area, extent, and/or quality of habitat. The main threats for this species are habitat loss and contamination due to cattle ranching, agriculture, and mining activities (see Discussion).





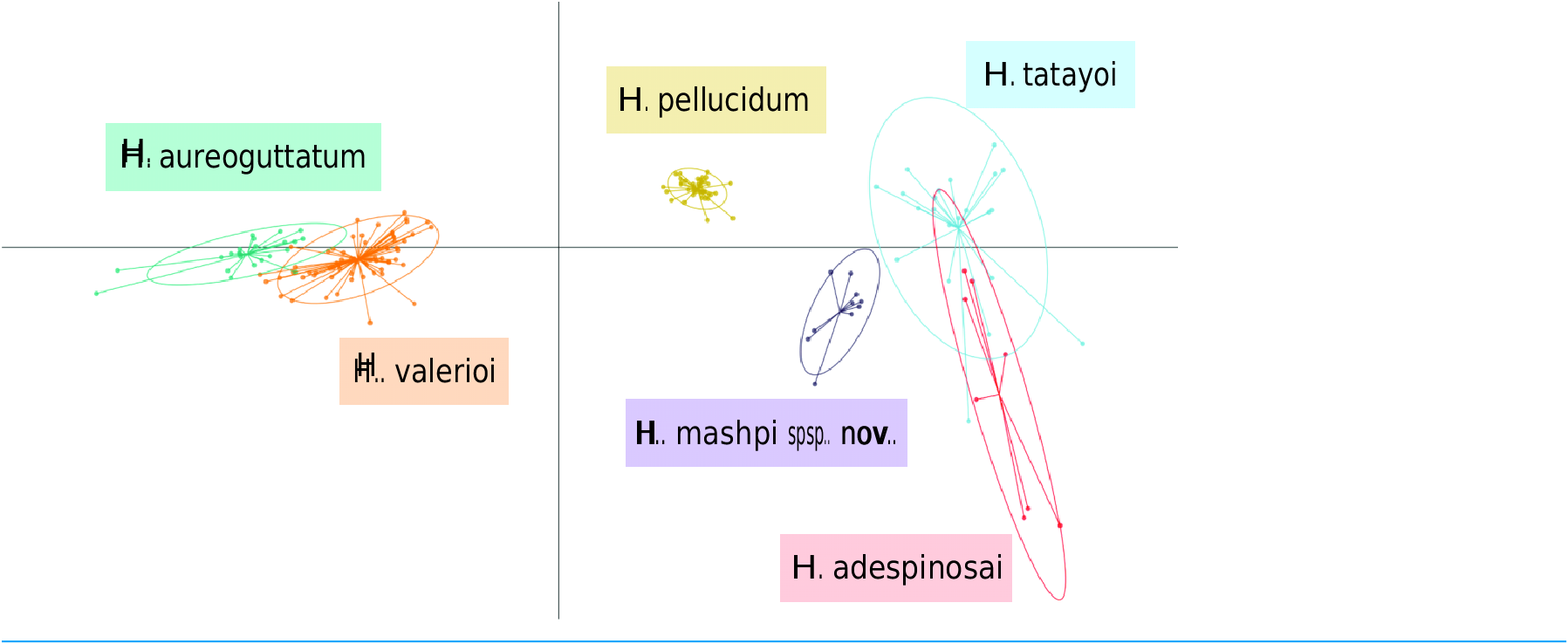



Table 1 Differences between Hyalinobatrachium mashpi sp. nov., H. nouns sp. nov., and similar and closely related species.
| Species | SVL in mm (adult males) | Dorsal pattern | Hear coloration in life | Interorbital bar | Type of call | Biogeographic distribution/ Country/ Elevation (m a. s.l.) | Sources |
|---|---|---|---|---|---|---|---|
| H. adespinosai | 20.5–22.2 | Pale yellowish green with small pale yellow spots and minute gray to black melanophores | Red | Absent | Pulsed | Amazonian slopes of the Andes Ecuador 1,670–1,795 | Guayasamin et al. (2019a) |
| H. anachoretus | 20.6–21.4 | Apple green with small yellow spots and minute melanophores | Red | Absent | Pulsed | Amazonian slopes of the Andes Peru 2,001–2,050 | Twomey, Delia & Castroviejo-Fisher (2014) |
| H. aureoguttatum | 20.4–24.0 | Greenish yellow with large, bright yellow spots and, in some populations, dark flecks | Usually red, but also red and white, or white | Usually absent | Tonal | Chocó, Pacific slopes of Andes Colombia, Ecuador, Panama 0–1,340 | Barrera-Rodriguez & Ruiz-Carranza (1989), Guayasamin et al. (2020), this study |
| H. bergeri | 20.3–22.4 | Apple green with small yellow spots and minute melanophores | Mostly white | Absent | Tonal | Amazonian lowlands and Amazonian slopes of the Andes Peru, Bolivia 300–1,980 | Castroviejo-Fisher et al. (2009) |
| H. chirripoi | 24–27 | Greenish yellow with small yellow spots | Red | Absent | Pulsed | Central America (Costa Rica, Panama); Chocó (Colombia and Ecuador) 0–320 | Savage (2002), Kubicki (2007), Guayasamin et al. (2020) |
| H. colymbiphyllum | 23–30 | Greenish yellow with yellow spots | Red | Absent | Pulsed | Central America (Costa Rica, Honduras, Panama); Chocó (Colombia) 0–1,710 | McCranie & Wilson (2002), Savage (2002), Kubicki (2007) |
| H. esmeralda | 18.4–22.3 | Greenish yellow with yellow spots and some minute dark dots | Red or red and white | Absent | Unknown | Amazonian slopes of the Andes Colombia 1,026–1,700 | Ruiz Carranza & Lynch (1998), Acosta-Galvis (2017), Twomey, Delia & Castroviejo-Fisher (2014) |
Table 2 Acoustic differences between Hyalinobatrachium mashpi sp. nov. and related species.
| Species | Call codes | Call structure | Call duration | Inter-call | Peak | Maximum | Minimum |
|---|---|---|---|---|---|---|---|
| (s) | interval (s) | frequency | frequency | frequency | |||
| (kHz) | (kHz) | (kHz) | |||||
| H. mashpi | LBE-C-051, LBE- | 1 note per call; pulsed; | 0.37–0.46 | 10.07–17.48 | 5.25–5.6 | 5.46–5.81 (5.62 | 4.62–4.92 (4.79 |
| sp. nov. | C-052 | 8–10 pulses per note | (0.43 ± 0.03) | (12.8 ± 2.17) | (5.38 ± | ± 0.11) | ± 0.10) |
| two individuals, | 0.12) | ||||||
| 12 calls | |||||||
| H. adespinosai | LBE-C-050 | 1 note per call; pulsed; | 0.448–0.646 | 10.87–30.04 | 4.87–5.04 | 5.0–5.25 | 4.32–4.75 |
| one individual | ~12 pulses per note | (0.543 ± | (20.12 ± 8.77) | (4.94 ± | (5.11 ± 0.08) | (4.57 ± 0.15) | |
| 10 calls | 0.07) | 0.07) | |||||
| H. aureoguttatum | LBE-C-053–057 | 1 note per call; tonal; | 0.078–0.087 | 1.99–5.20 | 6.63–7.41 | 7.78–8.90 (8.11 | 5.16–5.91 |
| six individuals | harmonics present | (0.082 ± | (3.24 ± 0.79) | (6.87 ± | ± 0.27) | (5.62 ± 0.21) | |
| 24 calls | 0.002) | 0.25) | |||||
| H. chirripoi | LBE-C-010 | 1 note per call; pulsed; | 0.235–0.274 | 84.3 | 4.48 | 4.99–5.77 (5.38 | 4.16–4.21 |
| one individual | 12–13 pulses per note | (0.255 ± | (only 2 notes | (4.48 ± 0) | ± 0.05) | (4.19 ± 0.04) | |
| two calls | 0.03) | in recording) | |||||
| H. pellucidum | LBE-C-003 | 1 note per call; tonal | 0.1–0.146 | 1.67–5.35 | 5.60–5.86 | 5.86–6.14 | 5.05–5.32 |
| 1 individual | (0.129 ± | (2.94 ± 0.79) | (5.70 ± | (6.0 ± 0.06) | (5.16 ± 0.07) | ||
| 41 calls | 0.009) | 0.06) | |||||
| H. tatayoi | LBE-C-058 | 1 note per call; tonal | 0.076–0.276 | 2.05–21.68 | 4.45–5.11 | 4.83–5.40 (5.14 | 3.30–4.61 (4.24 |
| four individuals | (0.143 ± | (7.64 ± 4.92) | (4.82 ± | ± 0.17) | ± 0.34) | ||
| 26 calls | 0.04) | 1.77) | |||||
| H. valerioi | ML 201469, | 1 note per call; tonal; | 0.065–0.10 | 1.76–8.00 | 6.46–7.24 | 7.22–7.90 | 4.09–5.88 (5.12 |
| three individuals | ML201473, | harmonics present | (0.079 ± | (4.27 ± 1.21) | (6.77 ± | (7.53 ± 0.17) | ± 0.51) |
| 70 calls | ML201475 | 0.01) | 0.19) |
Table 3 Results from the discriminant analysis of principle components (DAPC), comparing the advertisement calls of Hyalinobatrachium mashpi sp. nov. with those of closely related species (see Fig. 7).
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| PCA Eigenvalues | 23.76 | 1.48 | 0.16 |
| PCA Loadings | |||
| Peak Frequency | 0.08 | −0.57 | −0.09 |
| Call (Note) Duration | −0.02 | 0.023 | 0.05 |
| Inter-call Interval | −0.99 | −0.14 | 0.02 |
| Maximum Frequency | 0.11 | −0.78 | −0.19 |
| Minimum Frequency | 0.04 | −0.21 | 0.98 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |