
Gyrodactylus breviradix, Vega & Razzolini & Arbetman & Viozzi, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4664.3.9 |
|
publication LSID |
lsid:zoobank.org:pub:06BEFFC0-EB9E-4B77-97A0-7F3D497EA395 |
|
persistent identifier |
https://treatment.plazi.org/id/FB2E87F3-FFAB-4F51-FF00-DB8EF2278CBC |
|
treatment provided by |
Plazi |
|
scientific name |
Gyrodactylus breviradix |
| status |
sp. nov. |
Gyrodactylus breviradix View in CoL n. sp.
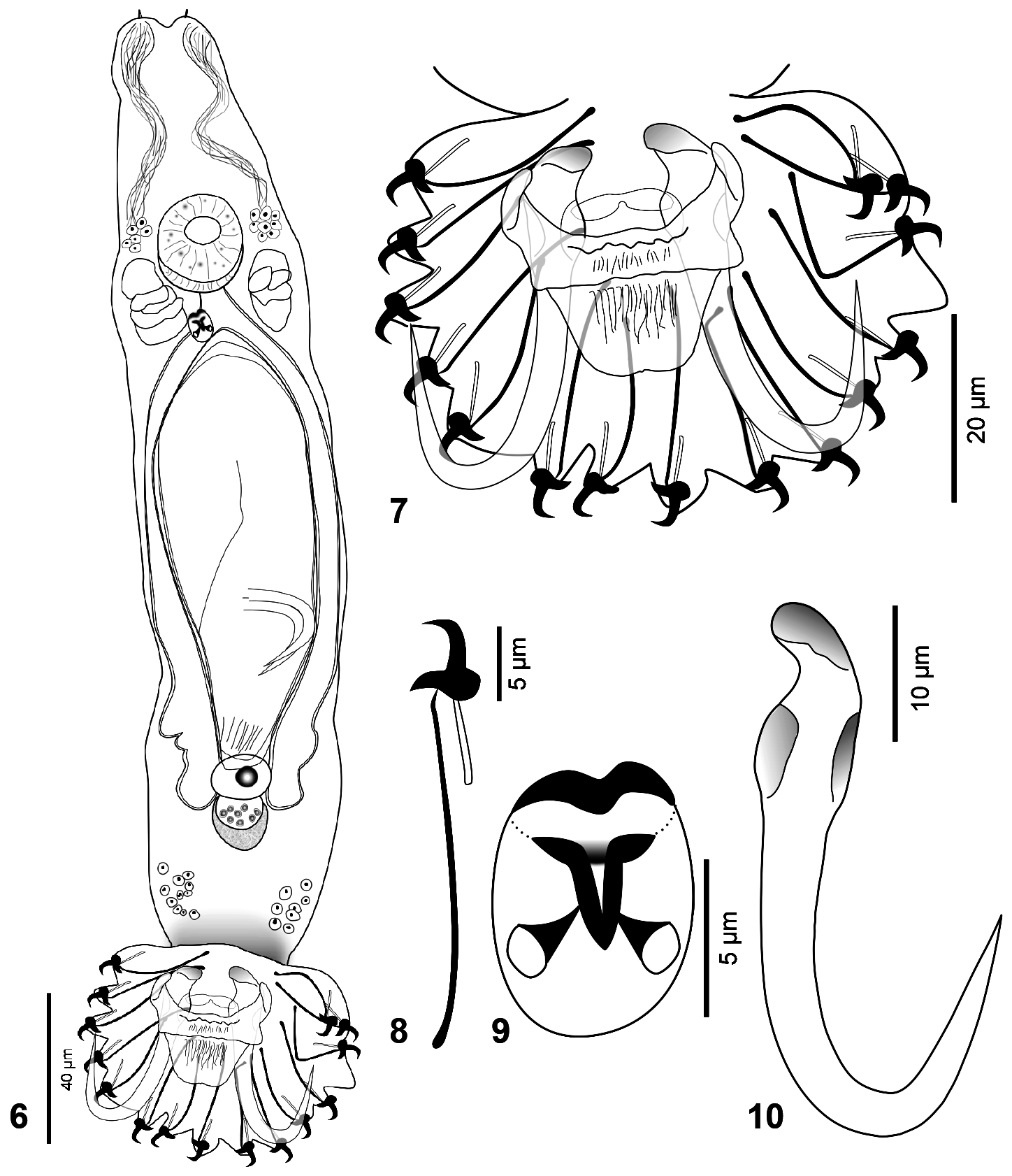
( Figs 6–10 View FIGURES 6–10 , 12 View FIGURES 11–12 , Table 1 View TABLE 1 )
urn:lsid:zoobank.org:act:8DE2A9EA-BDD7-4980-924E-AF36D14D416D
Type host: Cnesterodon decemmaculatus (Jenyns, 1842) ( Cyprinodontiformes : Poeciliidae ).
Type locality: Neuquen River , Vista Alegre, Neuquen, Argentina (38°59’36.6”S 67°59’31.1”W) GoogleMaps .
Site of infection: Body surface.
Specimen studied: holotype, MACN-Pa 679; paratypes MACN-Pa 680/1, MACN-Pa 680/2; hologenophore, MACN-Pa 681.
Specimen sequenced: ITS1 (GenBank accession numbers: MK 312262 View Materials , MK 312261 View Materials ), ITS2 (GenBank accession numbers: MK 312259 View Materials , MK 312260 View Materials ), COII (GenBank accession numbers: MN 066552 View Materials , MN 066553 View Materials )
ZooBank registration: The life Science Identifier ( LSID) Gyrodactylus breviradix n. sp. is urn:lsid:zoobank.org:act:8DE2A9EA-BDD7-4980-924E-AF36D14D416D .
Etymology: The specific epithet refers to the short superficial root of the anchor.
Description: (Based on 9 specimens) Body elongate, 318 (238–364; n = 8) long, 74 (56–98; n = 9) wide. Cephalic glands, head organs, spike sensilla conspicuous. Cephalic glands anterolateral, lateral, posterolateral to pharynx. Pharynx composed of two tandem bulbs. Distal pharyngeal bulb muscular 26 (22–30; n = 5) wide; digitiform projections of distal pharyngeal bulb not observed; proximal pharyngeal bulb glandular 23 (19–27; n = 5) wide. Male copulatory organ ( MCO) 10 (8–11; n = 3) wide, armed with one spine, one row of two large spinelets, each with wide, ovate base. Testis ovate, 20 (n = 1) wide, overlapped by posterior portion of germarium. Germarium ovate, 11 (10–12; n = 2) long, 15 (10–20; n = 2) wide. Uterus with up to 2 generations of embryos. Syncytial follicles overlapping and immediately posterior to distal portions of caecae. Anchor 41 (36–43; n = 8) long; straight, point recurved; deep root poorly developed, knob-like; short superficial root, inwardly and dorsally recurved. Deep bar with a median notch at posterior edge. Superficial bar 23 (22-25; n = 3) long, 25 (22–27; n = 3) wide, with two robust, elongate anterolateral projections, folded inward; trapezoidal shield. Hook, 27 (26–29; n = 7) long with straight shank. Hooklet 6 (5–6; n = 4) long; point short, 90º angled, ending before level of toe tip; short uniform shafts; heel round, prominent; toe pointed, depressed; shelf convex; FH loop about 1/4 of shank length.
Remarks: Gyrodactylus breviradix n. sp. resembles Gyrodactylus xalapensis Rubio-Godoy, Paladini , García- Vásquez & Shinn, 2010, Gyrodactylus mexicanus Mendoza-Palmero, Sereno-Uribe & Salgado-Maldonado, 2009 , and Gyrodactylus takoke García-Vásquez, Razo-Mendivil , & Rubio-Godoy, 2015, having certain similarities in hooklet morphology ( Figure 12 View FIGURES 11–12 ). Gyrodactylus xalapensis differs by having a circular to sub-square heel, a shorter, robust toe, and a slightly upward-curved hooklet point ( Rubio-Godoy et al., 2010). Additionally, G. xalapiensis has an MCO with four spinelets ( Rubio-Godoy et al., 2010). Gyrodactylus mexicanus differs by having hooks with a proximally-disrupted proximal ligament, a shorter hooklet shaft with a recurved point, and the superficial roots of anchors folded posteriorly with grooves ( Mendoza-Palmero et al., 2009). Gyrodactylus takoke differs from G. breviradix n. sp. by having a straight point of the hooklet facing downwards and ending at the toe limit, a smaller hooklet ( Table 1 View TABLE 1 ), and a superficial bar with a triangular shield ( García-Vásquez et al., 2015).
Gyrodactylus breviradix View in CoL n. sp. hooks also resemble G. carolinae View in CoL , Gyrodactylus lebiasinus An, Jara & Cone, 1991 View in CoL , and Gyrodactylus bimaculatus An, Jara & Cone, 1991 View in CoL which parasitises characiform fishes, and Gyrodactylus pimelodellus An, Jara & Cone, 1991 View in CoL which parasitises siluriform fishes. Gyrodactylus carolinae View in CoL differs by having a robust shaft and hooklet point, a recurved hooklet point facing downwards that extends beyond the toe, and a superficial bar with a subtriangular shield ( Boeger et al., 2014). Gyrodactylus lebiasinus View in CoL and G. bimaculatus View in CoL differ by having a long hooklet point that extends beyond the limit of the toe, and a squarish toe ( An et al., 1991). Additionally, G. bimaculatus View in CoL differs by having a subtriangular shield and an MCO armed with 6 spinelets ( An et al., 1991). Gyrodactylus pimelodellus View in CoL differs by having a rectangular toe, a depressed heel, an MCO armed with 6 spinnelets, and large, parallel superficial bar projections ( An et al., 1991).
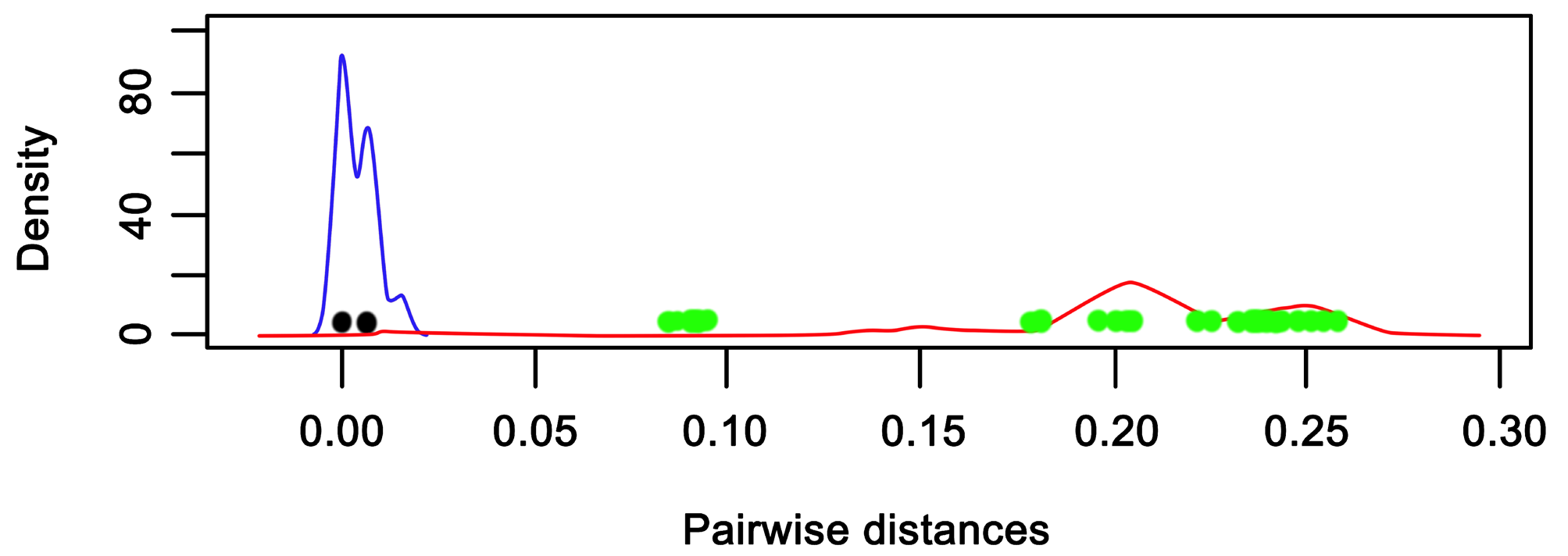
Molecular analysis. The new species described in this work were compared with species that were morphological and genetically similar. Not every species was available for each region of the ITS1 and ITS2, so we based our analyses mainly on the ITS2 region, where more comparisons were available. This was also true for the COII region, where none of the target species (i.e. more genetically or morphologically similar) were available. The genetic p-distances within and between species are shown in Table 2 View TABLE 2 . The trees recovered with both the Maximum Likelihood and the consensus Bayesian Inference tree showed identical topology for the ITS2 region.
Both sequences of G. decemmaculati specimens clustered in a single group showing no intraspecific distance ( Figure 13 View FIGURE 13 ). The interspecific genetic distances observed between the morphologically similar gyrodactylids from the cyprinodontiform fishes G. unami , G. turnbulli , and G. pictae were similar to those between most of the pairwise comparisons. Sequences of G. breviradix specimens also clustered in a single group and presented much shorter intraspecific distances (0.007) than those observed between the morphologically similar gyrodactylids from the cyprinodontiform fishes G. xalapensis (0.085) and G. takoke (0.092). However, the barcoding gap graph clearly separated G. decemmaculati from the other species, although for G. breviradix the p-distances to the smallest inter- specific distance were shorter. Nevertheless, the closest p-distance for G. breviradix was to G. takoke but this was still higher than the distance between G. takoke and G. xalapensis (0.010) ( Figure 14 View FIGURE 14 ).


| MK |
National Museum of Kenya |
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
| FH |
Fort Hays |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gyrodactylus breviradix
| Vega, Rocio, Razzolini, Emanuel, Arbetman, Marina & Viozzi, Gustavo 2019 |
Gyrodactylus breviradix
| Vega & Razzolini & Arbetman & Viozzi 2019 |
G. carolinae
| Boeger, Ferreira, Vianna & Patella 2014 |
Gyrodactylus carolinae
| Boeger, Ferreira, Vianna & Patella 2014 |
Gyrodactylus lebiasinus
| An, Jara & Cone 1991 |
Gyrodactylus bimaculatus
| An, Jara & Cone 1991 |
Gyrodactylus pimelodellus
| An, Jara & Cone 1991 |
Gyrodactylus lebiasinus
| An, Jara & Cone 1991 |
G. bimaculatus
| An, Jara & Cone 1991 |
G. bimaculatus
| An, Jara & Cone 1991 |
Gyrodactylus pimelodellus
| An, Jara & Cone 1991 |