
Glassella nataliae, Salgado-Barragán & Barragán-Zepeda, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4952.2.10 |
|
publication LSID |
lsid:zoobank.org:pub:07564B79-EB12-41D8-8BC8-FBED97BF6455 |
|
DOI |
https://doi.org/10.5281/zenodo.4694742 |
|
persistent identifier |
https://treatment.plazi.org/id/03C7E72E-FFC3-852B-FF68-C216FAE2F910 |
|
treatment provided by |
Plazi |
|
scientific name |
Glassella nataliae |
| status |
sp. nov. |
Glassella nataliae View in CoL new species
Material examined. 20 males and 21 females (including 9 ovigerous females), collected intertidally in six locations at Bahía Santa María-La Reforma lagoon, Sinaloa, Mexico; HOLOTYPE: male, ( CW 4.8 ; CL 2.4), La Garrapata Island, (25° 9’13.20”N, 108° 15’ 24.75”W), January 24, 2014, fine sand ( ICML-EMU 12523 ); GoogleMaps PARATYPES: La Garrapata Island , June 25, 2013 , 1 male, ( CW 4.5 ; CL 2.1) ( ICML-EMU 12524 A); January 24, 2014 , 2 males ( CW 5.4 – 5.5 ; CL 2.4–2.5), 2 OV ( CW 4.9 – 5.3 ; CL 2.3–2.4) ( ICML-EMU 12524 B); March 30, 2015 , 3 males ( CW 2.1–4.4; CL 1.0–2.2), 3 OV ( CW 5.3 –6.0; CL 2.5–2.7) ( ICML-EMU 12524 C); El Tambor , (25° 45’53”N, 108° 01’ 08”W), sand-mud, January 24, 2014, 1 female ( CW 5.6 ; CL 2.6) ( ICML-EMU 12525 A); January 18, 2015, 1 female ( CW 3.9 ; CL 1.9) ( ICML-EMU 12525 B); GoogleMaps Talchichilte Island (24° 56’27”N, 108° 02’ 55”W), fine sand, January 18, 2015, 1 male ( CW 4.8 ; CL 2.5), 5 females ( CW 4.4 – 5.3 ; CL 2.0–2.5) ( ICML-EMU 12526 ) GoogleMaps ; Saliaca Island , (25° 08’55”N, 108° 16’12”W), sandy mud, March 30, 2015, 7 males ( CW 3.2 – 4.3 ; CL 1.6–2.0), 2 females ( CW 3.8 – 4.2 ; CL 1.7–2.0), 1 OV ( CW 4.1 ; CL 1.9), ( ICML-EMU 12527 ) GoogleMaps ; same locality and date, 1 male ( CW 3.3 ; CL 1.6), 1 female ( CW 3.9 ; CL 1.9) (LEMA- CR-700) GoogleMaps ; 1 male ( CW 5.6 ; CL 2.5), 1 female ( CW 4.4 ; CL 2.1) ( CNCR 35490 ) ; Agua Dulce , (25° 02’12”N, 108° 13’34”W), sandy mud, March 30, 2015, 3 males ( CW 3.9 – 4.5 ; CL 1.9–2.3), 3 OV ( CW 3.8 ; CL 1.7) ( ICML-EMU 12528 ) GoogleMaps ; Meléndez Island (24°48’07”N, 108°03’22.3”W), sand, August 6, 2015, 1 female ( CW 7.0; CL 3.4) ( ICML-EMU 12529 ) GoogleMaps .
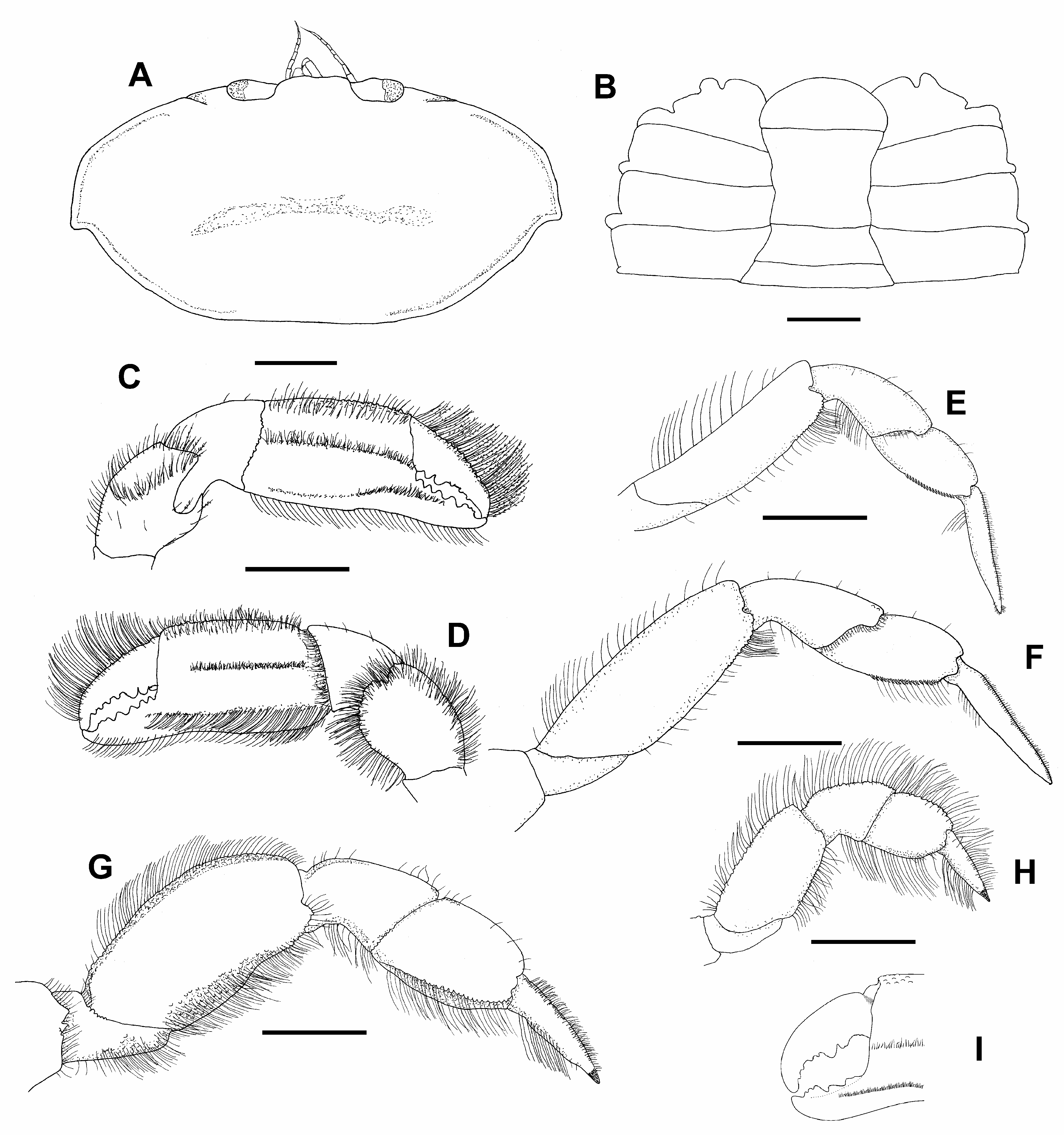
Description. Carapace ( Figs. 1A View FIGURE 1 , 2A View FIGURE 2 ) ovate, CW/CL = 1.7–2.1 in males (mean 2.05, n = 20); 1.9–2.3 in females (mean 2.15, n = 19), dorsally punctate, lacking transverse ridge; regions poorly marked, with transverse median depression and slightly elevated intestinal region. Anterolateral angles more convex than posterolateral; anterolateral ridge smooth, at most pitted, crossing branchial regions, falling short of orbit, extending to posterolateral margin; junction between anterolateral and posterolateral margins angled, directed posterolaterally.Anterior and lateral margins setose; front transverse, slightly produced and elevated anteriorly, with shallow median sulcus and incipient medial notch; orbits small, completely filled by eyes.
Fourth thoracic sternite with anterior and lateral notches, anterior the deepest ( Fig. 2B View FIGURE 2 ).
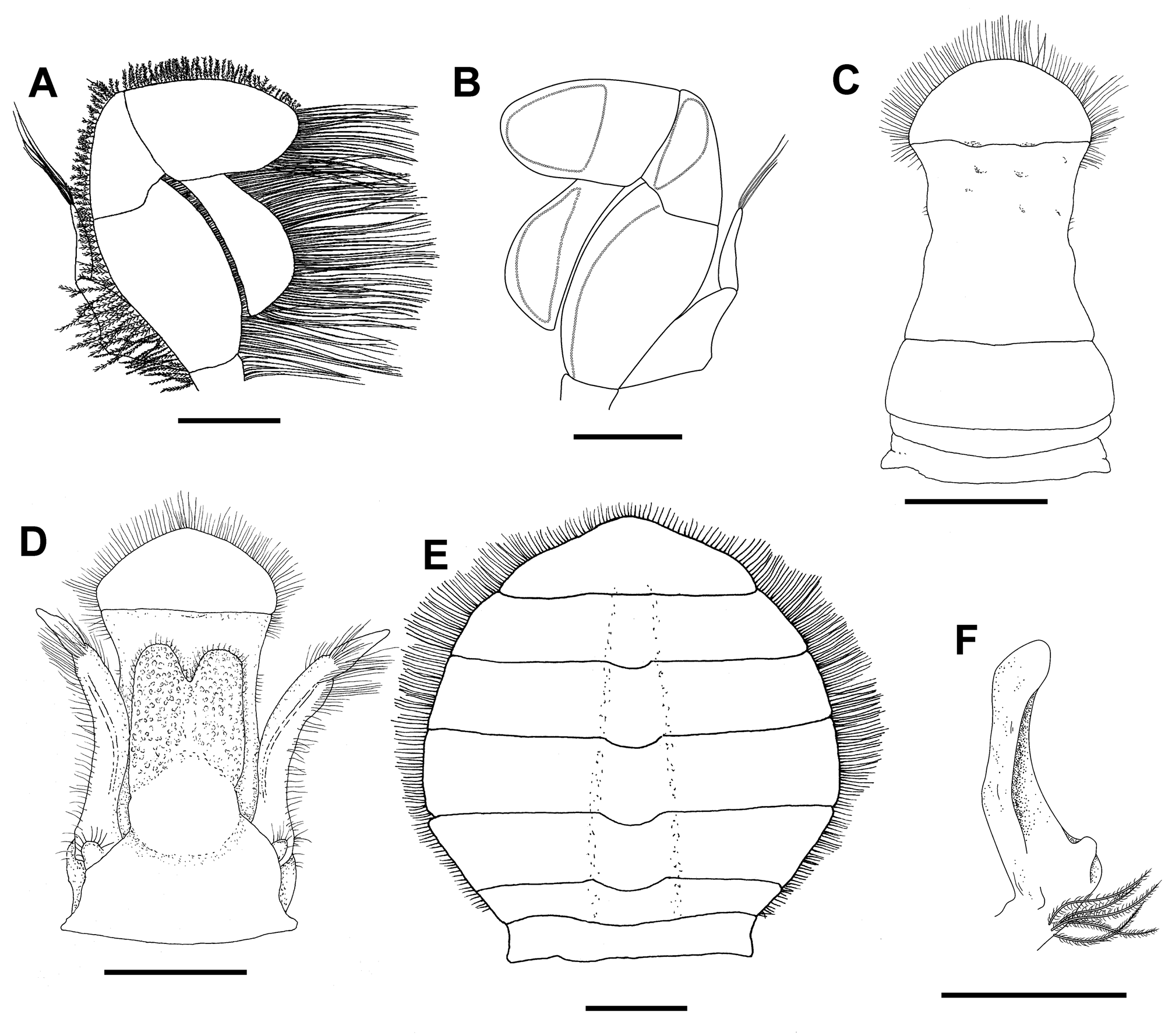
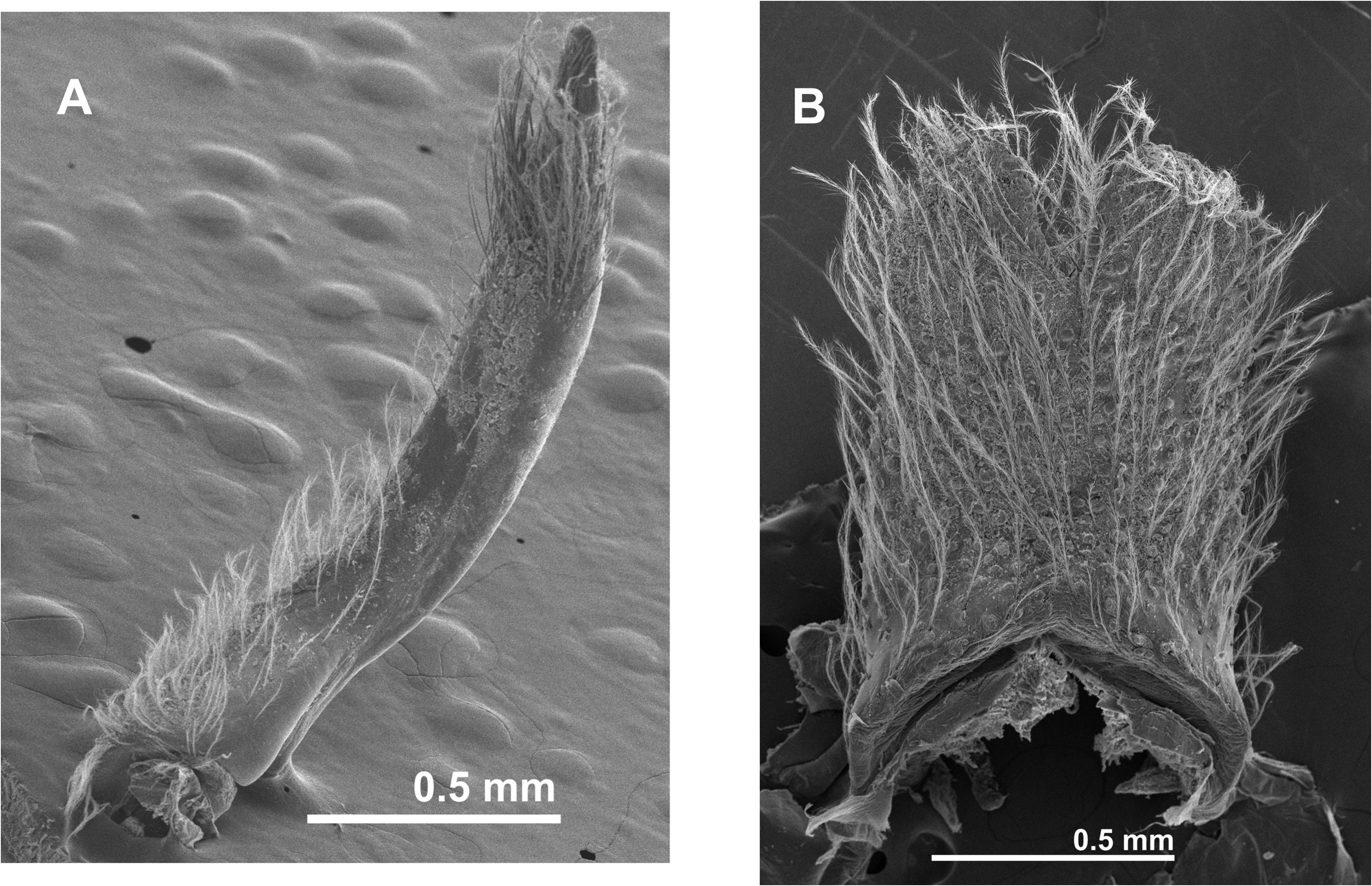
Third maxilliped ( Figs. 3A, B View FIGURE 3 ); ischium and merus fused (ischiomerus), trapezoidal, wider medially, lateral margins convex; carpus narrower than ischiomerus and shorter than propodus; propodus blunt, stout, inserted subdistally on carpus, laterally projected; dactylus about 0.8 as long as ischiomerus, inserted proximally on propodus, spatulated, broadened distally, mesial margin convex; setae in ischiomerus ( Figs. 3B View FIGURE 3 ) consisting of a simple row of long plumose setae on ventro-mesial margin and external margin with pappose setae; carpus, propodus and dactylus with sub-oval double-row pattern of long plumose setae emerging on mesial margin; exopodite two-segmented; basal segment with median protuberance in outer margin; distal segment slender, slightly curved, tapering, ending in long setae.
Chelipeds (Pereopods 1) ( Figs. 2C, D View FIGURE 2 ) stout, sub-equal, sub-cylindrical, similar in both sexes; merus slightly longer than carpus, dorsal surface almost flat, surrounded by long setae, ventral surface with semicircular protuberance, distomesial margin of protuberance granulated, surrounded by long setae; carpus short, with short setae on inner distal margin; palm about 1.5 times as long as dactylus, dorsum setose, with small granules on distal third, ventral margin slightly sinuous, inner surface ( Fig. 2D View FIGURE 2 ) with two longitudinal rows of setae, a median row of short setae from near proximal margin to the distal quarter of palm and subventral row of long setae from proximal margin to proximal part of pollex, setae longer proximally; outer surface ( Fig. 2C View FIGURE 2 ) with two longitudinal ridges of short setae medially and one short from the pollex to distal third and followed proximally by a shallow groove that continues to proximal margin; cutting edges of both movable and fixed fingers sub-parallel, tips deflected, pollex with 4–5 teeth, distal one bi or tricuspid; dorsum of dactyl convex, with sparse dorsal granules and dense long setae, cutting edge with 2 large, proximal teeth followed distally by 3–4 small teeth.
Pereopod 2 ( Fig. 2E View FIGURE 2 ) slender; merus long, about 3 times as long as wide, as long as carpus and propodus combined, flexor margin slightly convex, granulated distally, with sparse marginal setae and a tuft of long setae on distal margin, dorsal margin slightly concave, smooth, with sparse long setae; carpus longer than propodus, with sparse flexor setae; propodus ovate, flexor margin bicarinate, carina in posterior side distally serrate; dactylus slender longer than propodus, lined with dorsal setae on exterior margin, proximo-flexor with a tuft of longer setae.
Pereopod 3 ( Fig. 2F View FIGURE 2 ) similar but larger than Pereopod 2; merus about 2.5 times as long as wide, slightly longer than carpus and propodus combined, with sparse marginal setae and distal tuft of long setae on extensor margin, flexor margin distally granulated; carpus as long as propodus, extensor margin with distal sparce granules; flexor margin of propodus bicarinate, carinae distally serrated; dactylus slender, short setae on extensor and long setae on proximal flexor.
Pereopod 4 ( Fig. 2G View FIGURE 2 ) largest walking leg, stout; flexor margin of ischium with small tubercles, setose; merus 2 times as long as wide, shorter than carpus and propodus combined, flexor and extensor margins tuberculate, with long setae; carpus shorter than propodus; propodus with few tubercles on proximal extensor margin, flexor margin bicarinate, both carinae marginally serrate or denticulate, with long setae; dactylus as long as carpus, tapering, extensor margin with few proximal granules and short setae, flexor margin with long setae, tip corneous.
Pereopod 5 ( Fig. 2H View FIGURE 2 ) the smallest, stout, when extended, reaching to distal margin of merus of P4, extensor and flexor margins with long setae, extensor margin, from merus to propodus, granulate; merus subrectangular, about 2 times as long as wide, as long as carpus and propodus combined; carpus stout; propodus slightly longer than wide; dactylus as long as propodus, tip corneous.
Male pleon ( Figs. 3C, D View FIGURE 3 ) with somites 4–6 fused, telson and other somites free; first somite short, rectangular, second somite short, wider medially, third somite widest, lateral margin convex; fused somites 4–6 becoming narrower from proximal third somite to middle of fused somites and then wider to base of telson; telson distally rounded, slightly wider than somite 6 at base, distal part of lateral margins of somite 6 and margin of telson fringed with setae, longer and more abundant in telson.
Gonopodal plate of males ( Figs. 3D View FIGURE 3 , 4B View FIGURE 4 ) attached to ventral side of first pleonal somite, narrower and shorter than abdomen, lateral margins almost straight, distally bilobed, surface grainy, jelly-like texture and covered by long pappose setae; flanked by gonopods in situ.
Male first gonopods ( Figs. 3D View FIGURE 3 , 4A View FIGURE 4 ) curved, distally directed outward, fringed with long pappose setae, distal fifth constricted at base, terminating in acute angle and covered by setae. Male second gonopods ( Fig. 3F View FIGURE 3 ) slender, much smaller than first, external face concave, with long setae on basis.
Female pleon ( Fig. 3E View FIGURE 3 ) with 6 somites and telson, all free, broadly rounded in mature and ovigerous, not entirely covering sternites; first somite narrower in the middle; fourth somite widest; margins of pleonal somites and telson fringed with dense plumose setae, except for first somite.
Variations. The CW: CL ratio was similar between sexes; the mean ratio was slightly higher in females than in males (2.15 vs. 2.05). Chelae of the largest female (CW 7.0) are clearly unequal perhaps due to a process of regeneration, in addition, the occlusal margin of the dactyl in the large chela is more concave than in the rest of specimens, forming a gape with the pollex ( Fig. 2I View FIGURE 2 ). On the other hand, the pattern of number and shape of teeth in the cutting edges of the largest chela is the same as in the other specimens. Without considering abdominal differences between males and females, no other remarkable variations were observed between sexes. The presence of the GP was observed in all the males; that in the juvenile male (CW 2.1) appears much reduced.
Remarks. Although a genetic analysis of G. nataliae n. spec. could not be done, we include it into the genus Glassella on the basis of its morphological characteristics which clearly agree with the modified diagnosis for the genus Glassella made by Palacios Theil & Felder (2020) and slightly amended by Felder & Palacios Theil (2020) and that currently include four species previously included within the genus Pinnixa , the three species that had been placed in the genus Laminapinnixa and three new species recently described by Felder & Palacios Theil (2020) from the coasts of Florida, United States. Glassella nataliae n. spec. is very similar to G. miamiensis , an Atlantic species, by the general shape of the carapace, abdomen, pereopods and the third maxilliped, but they can be differentiated because the anterolateral carapaces ridges in the new species are smooth, not granulated, the male telson is rounded, not flattened and the GP has subparallel lateral margins and two distal lobes instead of convex margins and four distal lobes. Glassella faxoni differs from G. nataliae n. spec. by the shape of its chelae that forms a gape, and by having a triangular (vs. rounded) telson. In accordance with the description of G. vanderhorsti ( Rathbun, 1924, Plate III, figs. 7, 8, text-figs. 1 and 2), this species can be distinguished from G. nataliae n. spec. because the former has a tapered pleon, with undulating margins and an oblong telson, while in the new species the pleon has its narrowest part in the middle of the fused segments 4–6 with the last somite wider than preceding with no markedly undulating margins and the telson is more rounded. On the other hand, according to the figures provided by McDermott (2014) and Palacios Theil & Felder (2020), the new species can be distinguished from G. vanderhorsti because in the latter the GP is almost rounded distally with sharp lateral projections, while the GP of the new species is distally bilobed, with straight lateral margins.
The new species can be distinguished from G. martini Felder & Palacios Theil, 2020 and G. taylori Felder & Palacios Theil, 2020 by the male pleon outline. Male pleon of G. martini has an inverted chalice outiline and that of G. taylori is broadly triangular while the male pleon of G. nataliae n. spec. has an hourglass outline. On the other hand, G. martini has a carapace covered with fine granules and the anterolateral ridge of the carapace is slightly elevated while G. nataliae n. spec. has a dorsally glabrous carapace and a raised anterolateral ridge, not granulated. On the other hand, in G. taylori the anterolateral ridge of the carapace is granulated and the propodus and dactylus of the third maxilliped are short (vs. long in the new species). Glassella chaneyi Felder & Palacios Theil has a male pleon with hourglass outline, similar to that of the new species but in the former the GP is short, about 0.5 the length of the gonopod, and distally bifid while the new species has a rectangular and distally bilobed GP of about 3/4 the length of the gonopod. On the other hand, G. chaneyi has an anterolateral ridge of carapace marked by closely set tubercles and its chelae are weak, laterally compressed with fingers narrowly elongated and the new species has non slender chelae and the anterolateral ridge is elevated, non-tuberculate.
In this study no specimens of G. abbotti , (until now the only species of Glassella in the eastern Pacific) could be reviewed, and there is no a published description of its pleon or pereopods 2-5, but, according to the original description made by Glassell (1935: 13) and comments on the male GP in Palacios Theil & Felder (2020: 96), it can be inferred that the new species is different from G. abbotti because the former has a bare dorsum of carapace, while in G. abbotti the dorsum of carapace is pubescent towards the external margins. On the other hand, the male GP of G. nataliae is clearly rectangular and distally bilobed. While there is not much information about the GP of G. abbotti, Palacios Theil and Felder (2020) pointed out “we can state that a clear ramification of the male gonopodal plate is present in the eastern Pacific species G. abbotti ”, which indicates that G. nataliae and G. abbotti have different gonopodal plates.
There are a number of species distributed in the eastern Pacific and included in the genus Pinnixa that could also conform to the general morphology of the genus Glassella . These are P. transversalis H. Milne Edwards & Lucas, 1844 , P. pembertoni Glassell, 1935 , P. bahamondei Garth, 1957 , P. darwini, Garth, 1960 and P. hendrickxi Salgado-Barragán, 2015 . These species also share the fact that they all have the male pleon with at least three of the abdominal somites completely or partially fused as defined for the genus Indopinixa and also observed in most of the species of Glassella (see: Naruse & Maenosono, 2012; Salgado-Barragán, 2015).
When comparing with the species mentioned above, G. nataliae n. spec. can be easily distinguished from P. transversalis because the latter has a sharp transversal ridge on the posterior region of the carapace that is absent in the new species. In P. bahamondei and P. transversalis , the posterior margin of the merus and propodus of walking legs 3 and 4 are spiny, on the other hand their male pleon is wider at abdominal somite 3 and becomes narrower towards the end of somite 6, with a wider telson than segment 6, while in the new species posterior margin of the merus and propodus of walking legs 3 and 4 are, at most, granulated on the dorso-distal portion of the propodus and the abdominal segment 6 is wider than 5 and followed by a wider telson, giving the pleon an “hourglass” outline. The new species can also be separated from P. pembertoni because this species has granulated anterolateral ridges on carapace (vs. smooth ones in G. nataliae n. spec.) and a male abdomen with segment 6 narrower than segments 4 and 5, in the same way as is indicated for P. transversalis and P. bahamondei while in G. nataliae n. spec. it has an “hourglass” outline. Pinnixa darwini is easily distinguished from the new species because it has the chelae with arched dactyli, forming a gape with the pollex (not gaping in the new species) and because its male telson is triangular instead of rounded in G. nataliae n. spec. When comparing the new species with P. hendrickxi , the exopod of the third maxillped in P. hendrickxi is short, curved at the tip and non-segmented and the distal process of male first gonopod is thin and relatively large, about one third of the total gonopod length while in G. nataliae the exopod of the third maxilliped has a slender flagellum and the distal process of the male first gonopod is stout and short, of approximately one fourth of the gonopod length.
Among the diagnostic characteristics of the genus Glassella, Palacios Theil and Felder (2020) pointed out that “the nine species in the Glassella-Indopinnixa complex shared… relatively stout legs with dactyli shorter than the propodi (especially for P4 and P5…)”. This characteristic is not fully met by specimens of G. nataliae n. spec., since their pereopods 2 and 3 are slender and clearly have a dactylus longer than the propodus. Not so for pereopods 4 and 5. This characteristic seems to be shared by G. miamiensis , since, although none of us were able to review specimens of that species, the photographs provided by McDermott ( McDermott, 2014, Figs. 3A, B View FIGURE 3 ) show two specimens whose pereopods 2 and 3 appear to have a dactylus as long as or longer than the propodus. Unfortunately, the angle of the photographs does not help to confirm this. In any case, we consider that the inclusion of the new species within Glassella is still valid and that this variation could be an adaptation due to its lifestyle inside galleries of other invertebrates.
Color in life. Dorsum of carapace and proximal portions of pereopods orange; tips of chelae, distal segments of pereopods 2–5, and ventral region creamy-white ( Fig. 1 View FIGURE 1 ).
Habitat. Not well defined, but apparently associated with species of Neotrypaea spp. In muddy-sand lagoon bottoms.
Distribution. Known only from Bahía Santa María-La Reforma coastal lagoon.
Etymology. The species is named in honor of the late mother of the first author (Natalia Barragán).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |