
Lasioglossum (Dialictus) problematicum (Blüthgen, 1923)
|
publication ID |
https://doi.org/ 10.5852/ejt.2015.137 |
|
publication LSID |
lsid:zoobank.org:pub:0CB83C33-E198-4B4F-A8F6-96C59B066112 |
|
DOI |
https://doi.org/10.5281/zenodo.3794621 |
|
persistent identifier |
https://treatment.plazi.org/id/5A060122-0E06-FFEF-17C2-AF7487B7663E |
|
treatment provided by |
Carolina |
|
scientific name |
Lasioglossum (Dialictus) problematicum (Blüthgen, 1923) |
| status |
|
Lasioglossum (Dialictus) problematicum (Blüthgen, 1923) View in CoL
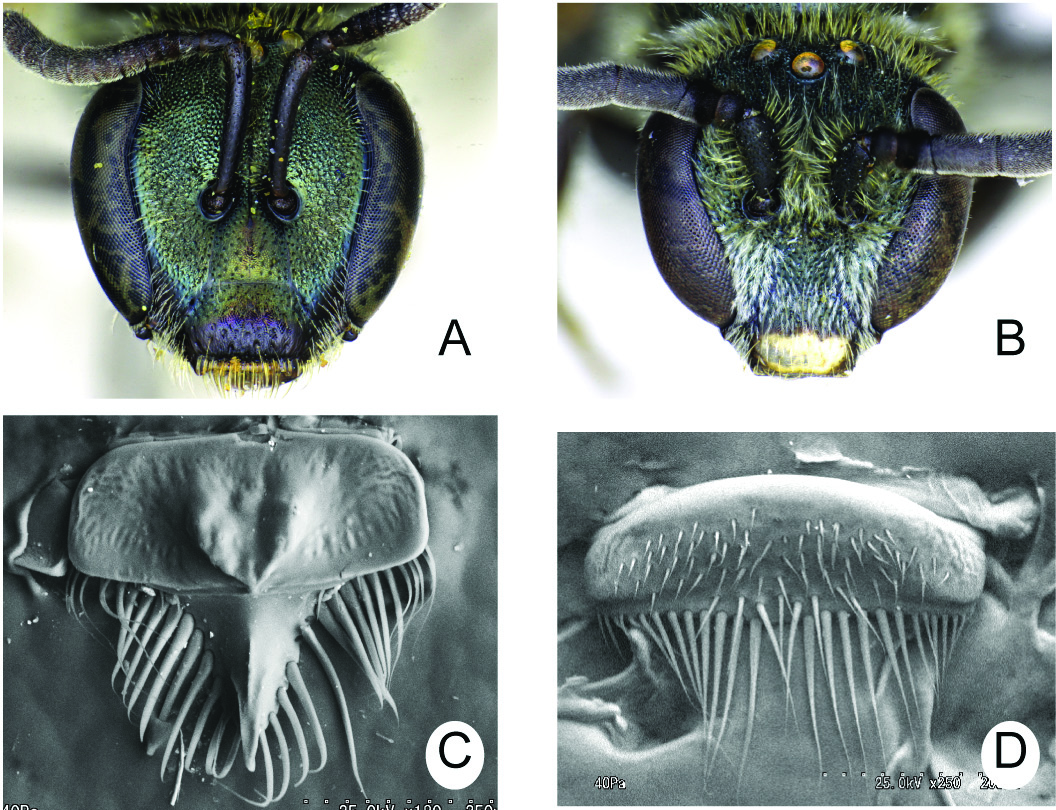
Figs 13A View Fig , 14 View Fig A–C, 15, 16, 19A, 27B
Halictus problematicus Blüthgen, 1923: 331 –332 (lectotype: Institute of Systematic and Experimental Zoology, Polish Academy of Sciences, Krakow, Poland, ♀; type locality: unknown (Siberia?); designated by Pesenko, 2007b: 115).
Halictus problematicus – Hirashima 1957: 15.
Lasioglossum (Evylaeus) problematicum View in CoL – Ebmer 1978: 311–312. — Ebmer 1996: 282; Ebmer 2002: 865–867 (♂). — Ebmer 2006: 563. — Murao et al. 2006: 50.
Evylaeus (Glauchalictus) problematicus – Pesenko 2007a: 26. — Pesenko 2007b: 85 (in key), 95 (in key), 115.
Diagnosis
This species is closely related to Lasioglossum alishanense Murao sp. nov., L. sanitarium (Blüthgen, 1926) and L. sichuanense Fan & Ebmer, 1992 from the northern Oriental Region, as stated above. For the differences between this species and L. alishanense , see the Key. It is also separated from the remaining two species by having the disc of the male S5 without an apical depression ( Fig. 14B View Fig ). The female cannot be clearly separated from the related species, as stated above.
Material examined
JAPAN: Hokkaido: 3 ♀♀, Rausu-seseki, Shiretoko, 16 Jun. 1984 (O. Tadauchi, ELKU); 3 ♂♂, Rebun Is., Rebun-no-oka, 7–8 Sep. 2005 (K. Goukon, cGou); 27 ♂♂, Kabuka, Rebun Is., 19 Aug. 1984 (O. Tadauchi, ELKU); 1 ♂, Air Port Wakkanai, 16 Aug. 1984 (O. Tadauchi, ELKU); 2 ♂♂, Botanical Garden, Sapporo, 3 Aug. 1959 (S. F. Sakagami & H. Fukuda, MNHAH); 1 ♂, Hamakoshimizu, 9 Aug. 1967 ( MNHAN); 10 ♂♂, Kami-Otoineppu, Nakagawa Exp. For., 14 Sep. 1969 (S. F. Sakagami & H. Fukuda, MNHAH); 1 ♂, Kitamoshiri, Hok. Uryu Exp. For., 14 Sep. 1969 (S. F. Sakagami & H. Fukuda, MNHAH); 2 ♂♂, Kussharo Lake, Teshikaga, 21 Jul. 1984 (O. Tadauchi, ELKU), 30 Aug. 1984 (O. Tadauchi, ELKU); 2 ♂♂, Nishiashoro, 9–10 Aug. 1955 (S. F. Sakagami & H. Fukuda, MNHAH); 1 ♂, Nissho Pass, Hidaka, 22 Sep. 1984 (O. Tadauchi, ELKU); 1 ♂, Cape Nosappu, Nemuro, 13 Sep. 1984 (O. Tadauchi, ELKU); 1 ♂, Nukabira, 10 Aug. 1965 (Y. Hirashima, ELKU); Sapporo, 1 ♂, 17 Aug. 1958, 4 ♂♂, 8 Sep. 1958 (S. Ueda, ELKU); 1 ♂, Shari Pass, Shari, 28 Jul. 1984 (O. Tadauchi, ELKU); 1 ♂, Shimamaki, Shiribetsu, 2 Sep. 1984 (O. Tadauchi, ELKU); 6 ♂♂, Shotoshibetsu, 20 Jul. 1984, 9 & 26 Aug. 1984 (O. Tadauchi, ELKU); 29 ♂♂, Sugatami, 6 Sep. 1967 ( MNHAH); 2 ♂♂, Tenneru, north Kushiro, 1968 (E. Ohtsuka, MNHAH); 4 ♂♂, Tokachimitsumata, Kamishihoro, 27 Aug. 1984, 11 Sep. 1984 (O. Tadauchi, ELKU); 6 ♂♂, Rubetu, Etorofu Is., 31 Aug. 1940 (S. Kuwayama & Y. Sugihara, ELKU); 13 ♂♂, Sibetoru, Etorofu Is., 6–7 Sep. 1940 (S. Kuwayama & Y. Sugihara, ELKU). Honshu: 4 ♂♂, Kuzukawa (Hiraka), Aomori Pref., 20 & 26 Aug. 1983 (M. Yamada, MNHAH); 1 ♂, Minami- Hakkoda (Hiraka), Aomori Pref., 17 Aug. 1984 (M. Yamada, MNHAH); 6 ♂♂, Mt. Iwaki, Aomori Pref., 7 & 21 Sep. 1980, 29 Aug. 1981 (M. Yamada, MNHAH); 4 ♂♂, Sukayu (Towadako), Aomori Pref., 2 Aug. 1987 (M. Yamada, MNHAH); 1 ♂, Takinosawa, Aomori Pref., 4 Sep. 1982 (M. Yamada, MNHAH); 1 ♂, Hachimantai, Akita Pref., 28 Jul. 1961 (M. Shiga, ELKU); 6 ♂♂, Mt. Akitakoma, Akita Pref., 28 Aug. 1972 (M. Honda, ELKU); 1 ♂, Mt. Chôkai-san, Akita Pref., 27 Aug. 1972 (M. Honda, ELKU); 1 ♂, Fukenoyu spa, Kazono city, Akita Pref., 2 Aug. 1974 ( T. & H. Suda, ELKU); 2 ♂♂, Mt. Zao, Shibakusadaira, Miyagi Pref., 27 Aug. 2004 (K. Goukon, cGou); 1 ♂, Mt. Zao, alt. 1300 m, Miyagi Pref., 28 Sep. 1996 (K. Goukon, cGou); 2 ♂♂, Mt. Zao, Miyagi Pref., 31 Aug. 1979 (K. Goukon, cGou), 24 Aug. 1979 (Y. Maeta, ELKU); 15 ♂♂, Kazawa, Nagano Pref., 17 Aug. 1972 (Y. Yoshiyasu, ELKU); 2 ♂♂, Shiga Highlands, Nagano Pref., 16 Aug. 1972 (Y. Yoshiyasu, ELKU); 2 ♂♂, Sugadaira Highlands, Sanada-machi, Nagano Pref., 22 Aug. 2002 (K. Mitai, cMur); 2 ♂♂, Asama, Karuizawa, Nagano Pref., 23 Aug. 1967 ( T. & H. Suda, ELKU); 1 ♂, Shirakaba Lake, Chino city, Nagano Pref., 7 Sep. 1966 ( T. & H. Suda, ELKU); 1 ♂, Arimine, Ôyama-machi, Toyama Pref., 11 Aug. 1994 (H. Negoro, MNHAH); 39 ♂♂, Migatahara, Mt. Tate-yama, Toyama Pref., 25 Aug. 2004 ( T. Sugimoto, cMur); 1 ♂, Mt. Daicho, Katsuyama-shi, Toyama Pref., 27 Aug. 2004 ( T. Sugimoto, cMur); 1 ♂, Sen-ninmori, near Mt. Ikenodaira, north Alps, Toyama Pref., 19 Aug. 2005 (H. Hirano, cMur); 1 ♂, Tateyama-Migatahara, Toyama Pref., 16 Oct. 1995 (H. Negoro, MNHAH); 2 ♂♂, Mt. Haku-san, Ishikawa Pref., 6 Aug. 1970 (K. Kanmiya, ELKU), 9 Aug. 1974 (I. Togashi, ELKU); 1 ♂, Mt. Haku-san, alt. 1750– 1500 m, Ishikawa Pref., 22 Aug. 1988 (I. Togashi, ELKU).
Additional description
LABRUM ( Fig. 15 View Fig C–D). Basal area approximately 2.3 × as wide as long in female, 3 × in male; basal elevation of female moderately developed, that of male absent; distal process of female slender, nearly as long as basal area, and without lateral projection, that of male absent; keel of distal process narrow, apically pointed; labral fimbria acutely pointed at apex in both sexes.
STERNA. S4 medially with shallow groove on apical half; apical margin nearly straight. S5 without apical depression, medially weakly swollen and apico-medially cleaved; apical margin gently incurved. S6 normally shaped, not modified. S7–S8 ( Fig. 16F View Fig ): S7 with elongate, apically rounded median process, apex exceeding S8; S8 without median process.
MALE GENITALIA ( Fig. 16 View Fig A–E). Gonobase nearly flat at bottom, ventral arms connected with each other at upper ends; gonocoxite smooth, gently sloped in lateral view, and inner dorsal margin gently angulate at approximately basal 1/3; gonostylus large, located at top of gonocoxite, with dense short setae on inner surface; ventral retrorse lobe moderately long but not reaching gonobase, with dense short setae ventrally and relatively long blunt setae laterally; penis valve higher than gonocoxite, with high cleft on top.
Distribution
Russian Far East (Sakhalin) and Japan (Hokkaido, northern to central Honshu). Ebmer (1978, 2006) recorded this species from Primorsky and North Korea, respectively, based on female specimens. These records are not included in this paper, because the female cannot be clearly separated from that of some of the related species.
Flight period
Female: May to September in Hokkaido, Japan.
Male: August to October.
The flight record of females is based on specimens collected from the eastern part of Hokkaido, Japan. This record can be considered accurate, because one of the related species, L. virideglaucum Ebmer & Sakagami , does not sympatrically inhabit this area.
Flower records
In Japan this species visits the following 19 species in 6 families: Apiaceae : Angelica ursina (Rupr.) Maxim. Asteraceae : Anaphalis margaritacea (L.) Benth. & Hook. f. subsp. margaritacea ; Aster glehnii F. Schmidt ; Cirsium setosum (Willd.) M. Bieb. ; Crepidiastrum denticulatum (Houtt.) J.H. Pak & Kawano ; Erigeron annuus (L.) Pers.; Hieracium umbellatum L.; Picris hieracioides subsp. japonica (Thunb.) Krylov ; Rudbeckia laciniata L.; Senecio pseudoarnica Less. ; Solidago altissima L.; S. virgaurea L. subsp. asiatica (Nakai ex H. Hara) Kitam. ex H. Hara var. asiatica Nakai ex H. Hara ; S. virgaulea L. subsp. leiocarpa (Benth.) Hultén ; Sonchus brachyotus DC.; Taraxacum sp. Brassicaceae : Barbarea orthoceras Ledeb. Geraniaceae : Geranium thunbergii Siebold ex Lindl. & Paxton. Lamiaceae : Isodon sp. Rosaceae : Sorbaria sorbifolia (L.) A. Braun.
Biological reference
According to Sakagami & Kuribayashi (1979) and Sakagami et al. (1984), some biological information is available. Nest place and structure: the female prefers to make a nest at the forest edge; the nest structure is the IIIa type of Sakagami & Michener (1962). Social structure and life cycle: the brooding period of the overwintered generation is from early May to late July, in Sapporo city, Hokkaido; most of the overwintering females stay communally in the natal nests; the nest are communally reused in the next brooding season, hence many colonies are semisocial or sometimes delayed eusocial from the beginning of spring activities; females living in the same nests are mostly of the same generation and seem to be sisters in most cases, although some nests are inhabited by a two-year-old female and her daughters; all cohabiting females are inseminated, but only one of them has fully developed ovaries at the peak of brooding season, the others with undeveloped ovaries participate in foraging.



| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lasioglossum (Dialictus) problematicum (Blüthgen, 1923)
| Murao, Ryuki, Tadauchi, Osamu & Lee, Heung-Sik 2015 |
Halictus problematicus Blüthgen, 1923: 331 –332
| Pesenko Y. A. 2007: 115 |
Evylaeus (Glauchalictus) problematicus
| Pesenko Y. A. 2007: 26 |
| Pesenko Y. A. 2007: 85 |
Lasioglossum (Evylaeus) problematicum
| Ebmer A. W. 2006: 563 |
| Murao R. & Ebmer A. W. & Tadauchi O. 2006: 50 |
| Ebmer A. W. 2002: 865 |
| Ebmer A. W. 1996: 282 |
| Ebmer A. W. 1978: 311 |
Halictus problematicus
| Hirashima Y. 1957: 15 |