
Evansula cumbraensis, Huys & Conroy-Dalton, 2006
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2006.00227.x |
|
persistent identifier |
https://treatment.plazi.org/id/03F487A0-FFB7-FF87-FFFE-FD2EFBB29635 |
|
treatment provided by |
Felipe |
|
scientific name |
Evansula cumbraensis |
| status |
sp. nov. |
EVANSULA CUMBRAENSIS SP. NOV.
Synonym: E. pygmaea (T. Scott, 1903) sensu Mielke (1975) .
Original description: Mielke (1975: 98–100, abb. 66, as E. pygmaea : figs 2b, 3a–c).
Additional description: Martínez Arbizu & Moura (1994) (as E. pygmaea ).
Type locality: Scotland, Isle of Cumbrae , Fintray Bay; exposed sandy beach with a steep profile; coarse sand .
Material examined: (1) From type locality: holotype ♀ ( NHM reg. no. 2005.2025) and paratype ♂ ( NHM reg. no. 2005.2026) dissected on six and five slides, respectively; other paratypes in alcohol 15 ♀♀ and 7 ♂♂ ( NHM reg. nos 2005.2027−48); coll. R. Huys & S. Conroy-Dalton; 27 August 1993; Karaman- Chappuis method ( Delamare Deboutteville, 1954); (2) Firth of Forth , Elie Bay ( Scotland): 15 ♀♀ and 10 ♂♂ in alcohol; coll. R. Huys & S. Conroy-Dalton; 26 August 1993; sandbank sampled at low tide; clean medium sand; deposited in NHM (reg. nos 2005.2049− 58); (3) From Dr W. Mielke: Isle of Sylt; 5 ♀♀ and 5 ♂♂ in alcohol ( NHM reg. nos 2005.2059−68) .
Etymology: The species is named after the type locality, the Isle of Cumbrae in Scotland.
Description
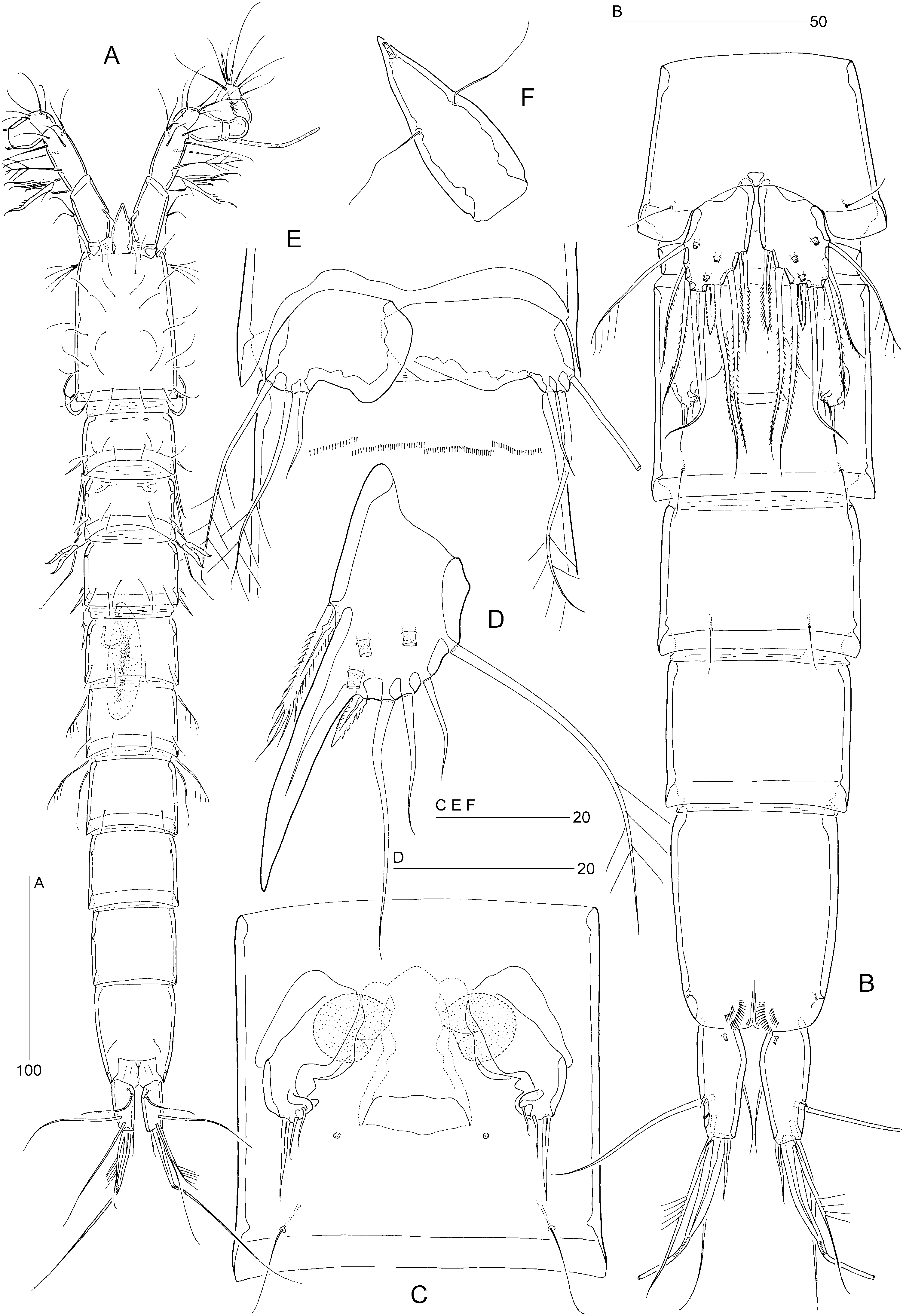
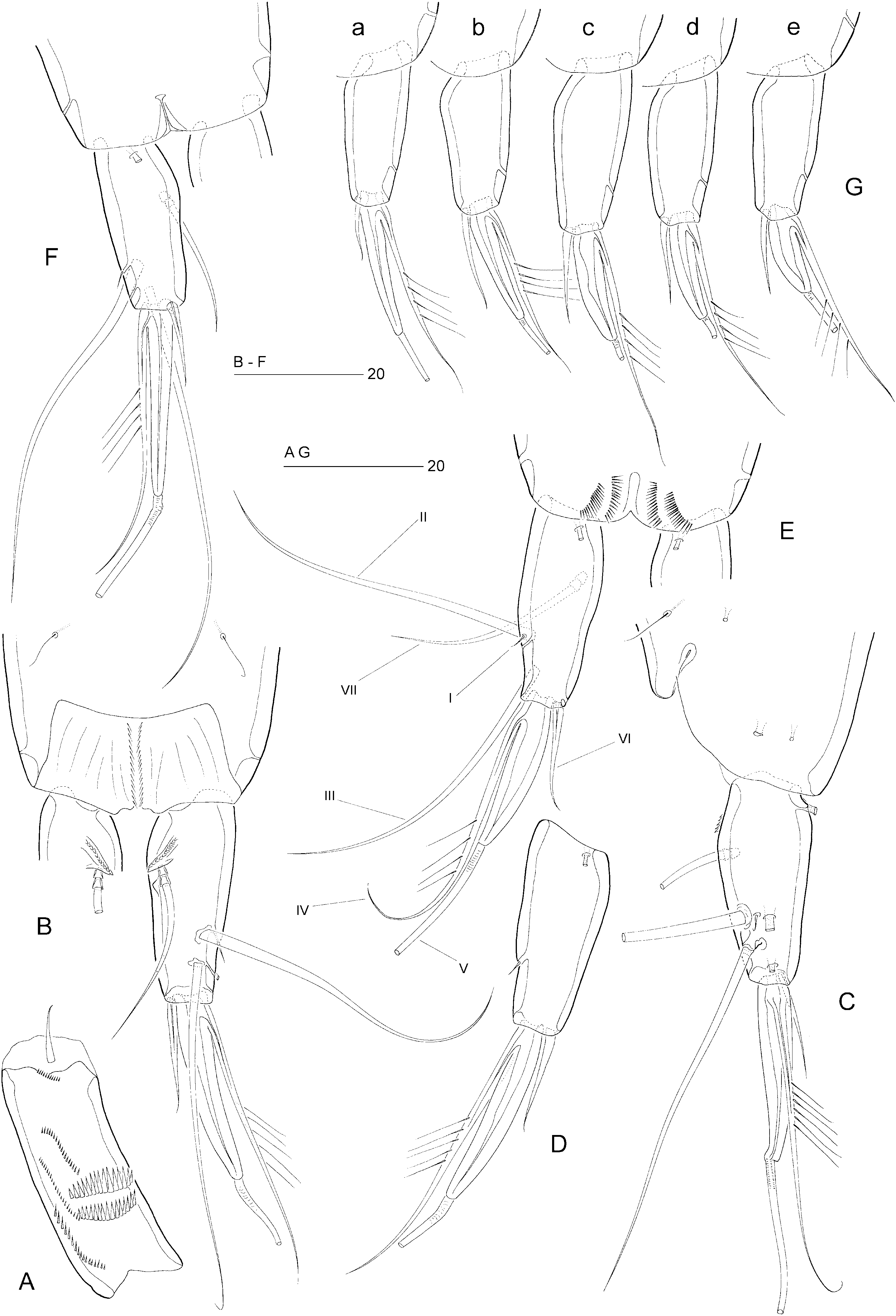
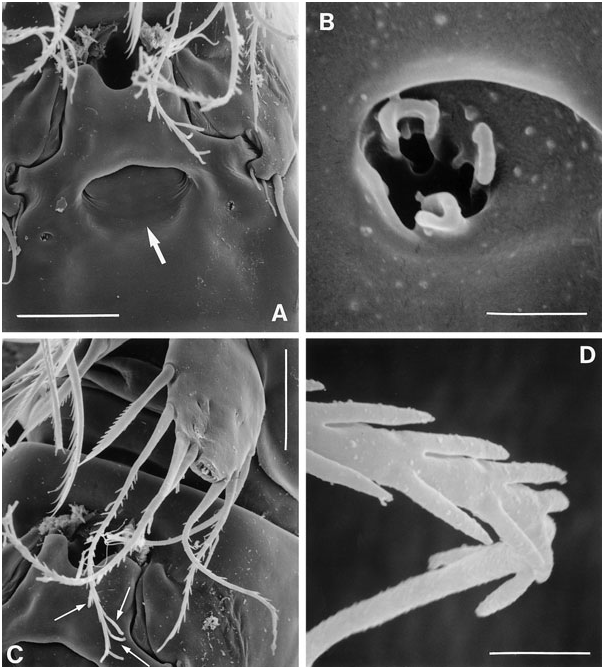
Female: Total body length: 500–560 µm (N = 10; mean = 535 µm). Body slender, cylindrical, semitransparent, colourless; no distinct separation between prosome and urosome. Genital double-somite completely fused ( Fig. 18B, C View Figure 18 ); original segmentation marked dorsally by paired anterior and posterior sensillae. Anal somite longer than wide (60 × 44 µm), with three pairs of secretory pores laterally ( Fig. 21C View Figure 21 ); posterior margin with two short spinular rows on either side of ventral midline ( Figs 18B View Figure 18 , 21E View Figure 21 ). Anal operculum weakly developed, unarmed ( Fig. 21B View Figure 21 ).
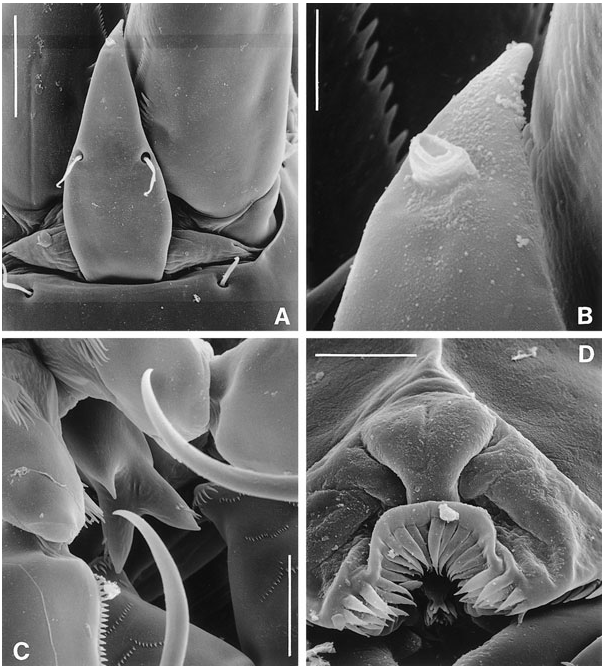
Caudal ramus slightly conical ( Figs 18B View Figure 18 , 21B–E View Figure 21 ), length (measured along outer margin) approximately 2.5 times the proximal width; dorsal surface without chitinous spur but with raised spinular row forming crest covering base of seta VII ( Figs 21B View Figure 21 , 27C, D View Figure 27 ); with seven setae, setae I–VI in distal and seta VII in proximal half ( Fig. 21B View Figure 21 ); seta I diminutive, tubular ( Fig. 27A, B View Figure 27 ); setae II–III long and bare; seta IV long, longer than styliform part of V, plumose half its length; seta V long, with distinct flexure zone between styliform basal part and long distal flagellate part, fused at base with seta IV ( Fig. 27A View Figure 27 ); seta VI vestigial; seta VII tri-articulate at base ( Fig. 27D View Figure 27 ) and located along proximal inner margin; ventral surface with one tube-pore near anterior margin ( Fig. 21E View Figure 21 ), lateral surface with two tube-pores ( Figs 21C View Figure 21 , 27A, B View Figure 27 ).
Rostrum elongate ( Figs 18F View Figure 18 , 23A, B View Figure 23 ), with parallel margins in proximal half, tapering distally; distinctly shorter than first antennulary segment; base surrounded by area of flexible integument ( Fig. 23A View Figure 23 ), with two long sensillae; median pore raised and positioned dorsally near apex of rostrum ( Fig. 23A, B View Figure 23 ).
Antennule seven-segmented; armature formula as in E. incerta ; segment 1 only slightly shorter than segment 2, with small sclerite around proximal posterior margin ( Fig. 23A View Figure 23 ), with pattern of spinular rows as illustrated in Figure 21A View Figure 21 .
Antenna, mandible, maxillule, and maxilla as in E. incerta .
Labrum, a massive lobe ( Fig. 22A View Figure 22 ) overlying mouth and mandibular gnathobases; with series of overlapping spinular rows around midventral margin ( Figs 22C View Figure 22 , 23D View Figure 23 ).
Paragnaths, well-developed lobes, closely adpressed to gnathobases of mandibles ( Fig. 22C View Figure 22 ); with two rows of spinules each.
Maxilliped ( Fig. 22A View Figure 22 ) as in E. pygmaea . Syncoxa without seta. Palmar margin of basis with spinular row posteriorly and patch of longer setules anteriorly. Endopodal claw distinctly curved, bare. Area between maxillipedal syncoxae and intercoxal sclerite of P1 provided with midventral, trifid, spinous process ( Figs 22A View Figure 22 , 23C View Figure 23 ).
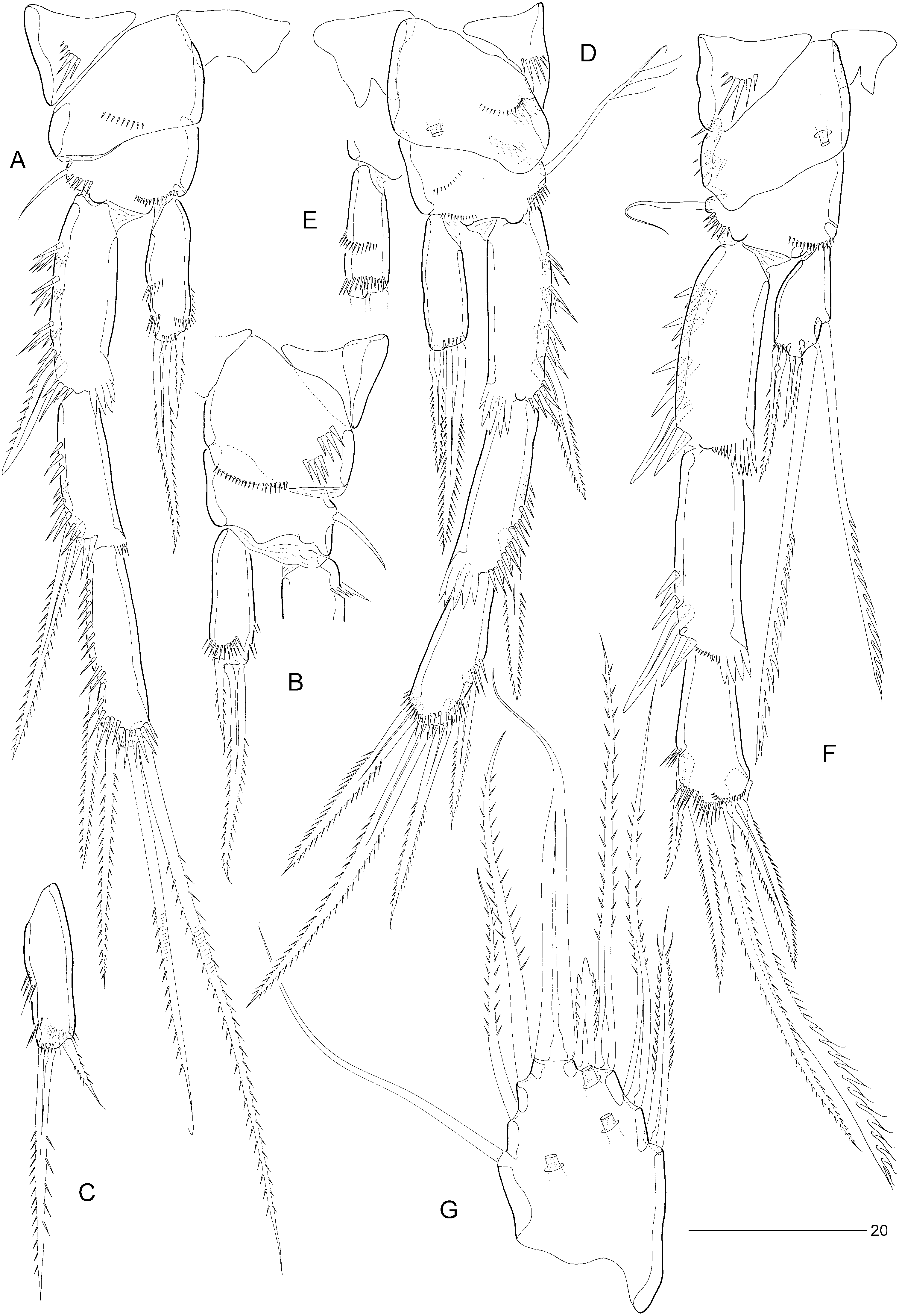
P1 ( Fig. 19A, B View Figure 19 ). Praecoxa strongly developed. Coxa with three distinct spinular rows. However, scanning electron microscopy ( Fig. 23C View Figure 23 ) revealed additional rows of tiny spinules, which cannot be discerned using differential interference contrast microscopes. This additional ornamentation was observed in E. arenicola and it is probable that the same spinular pattern is present in all species. Basis with long, naked, inner seta and short, pinnate, outer seta. Exopod three-segmented; with two spines and two geniculate setae on exp-3. P1 endopod prehensile, distinctly longer than exopod; proximal segment approximately ten times as long as average width, with pinnate inner seta being plumose in proximal third; distal segment short, with two spinular rows, a subdistal setule, and two short, geniculate spines distally.
Swimming legs P2–P4 ( Fig. 20A–F View Figure 20 ). P4 distinctly longer than P2–P3. Width of intercoxal sclerites decreasing in antero-posterior direction ( Fig. 20A, D–F View Figure 20 ). Praecoxae well developed, with spinular row on anterior surface. Coxae with pattern of spinules as in Figures 20A, B, D–F View Figure 20 , with a large tube-pore on anterior surface of P3 and P4. Bases with outer seta (short and bare in P2, long and plumose in P3, long and bare in P4); with spinular rows on anterior surface only ( Fig. 20A, B, D–F View Figure 20 ). Exopods three-segmented, endopods one-segmented. Exopodal spines of P2 exp-2 and exp-3 elongate. Inner distal spine of P3–P4 exp-3 shorter than outer distal one. Inner setae of P4 endopod and exp-3 serrate. Inner element of P2 endopod spiniform, pinnate, less than half length of distal spine. Armature elements of P3 endopod spiniform; inner spine longer than half length of outer spine. Seta and spine formulae as for genus.
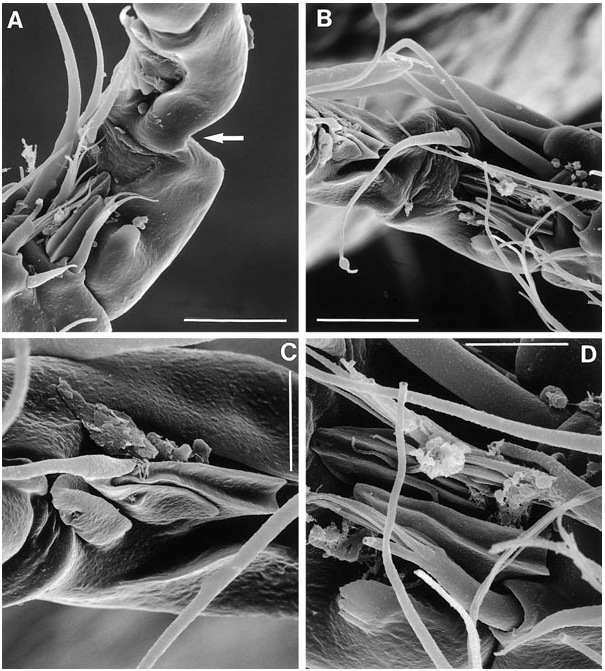
Fifth pair of legs ( Figs 18B View Figure 18 , 20G View Figure 20 ) with baseoendopod and exopod fused into a common elongate plate, tapering distally, with stepped inner and outer margins; apex with strong, articulating spine, distinctly longer than plate and with flagellate tip; outer margin with one bare (seta derived from baseoendopod) and two pinnate setae; inner margin with two serrate spines and two long, pinnate setae (all free at base); anterior surface with three large tube-pores.
Sixth legs ( Figs 18C View Figure 18 , 26A View Figure 26 ) each represented by small operculum closing off gonopore; armature consisting of spiniform outer element and two accessory setules. Genital apertures not fused medially; copulatory pore very wide, located at level of gonopores ( Fig. 26A View Figure 26 ); leading via short chitinized copulatory duct to paired anterior extensions positioned anterior to genital apertures; copulatory pore flanked by two complex secretory pores, each consisting of four vents ( Fig. 26B View Figure 26 ).
Male: Body length: 480–525 µm (N = 5; mean = 505 µm) ( Fig. 18A View Figure 18 ). Spermatophore 65 µm. Anal somite without spinules on ventral anterior and hind margins ( Figs 21F View Figure 21 , 25D View Figure 25 ).
Antennule indistinctly nine-segmented; geniculation between segments 7 and 8 ( Fig. 24A View Figure 24 ); segment 1 with one minute seta and spinular pattern as in female; segment 2 longest, with one plumose and eight naked setae; segment 3 with six setae; segment 4 minute, forming an incomplete ring, with two short setae; segment 5 distinctly swollen, with six elements along the anterior margin and with a distal cylindrical process bearing a basally fused seta and large, constricted aesthetasc; segment 6 with one long seta and one short pinnate spine; segment 7 with three spiniform elements and one seta ( Fig. 24D View Figure 24 ); segment 8 with one seta and three interdigitating modified setae ( Fig. 24B, C View Figure 24 ); segment 9 with five posterior setae, one anterior seta and one seta plus an acrothek apically. Apical acrothek consisting of two naked setae and one slender aesthetasc.
P2 ( Fig. 19D, E View Figure 19 ) with inner distal corner of basis not modified into spinous process. Exp-3 modified; inner distal element transformed into slender claw, directed medially and posteriorly, and with pinnules in distal half ( Fig. 19E View Figure 19 ). Endopod of approximately the same size as in female; outer margin with two spinular rows; apical seta longer and more slender, extending beyond distal margin of exp-2; inner seta minute, bare.
P3 ( Fig. 19F View Figure 19 ) with reduced hyaline frill on exp-1 ( Fig. 25B View Figure 25 ); exp-3 with secretory pore on anterior surface near joint with exp-2 ( Fig. 25A View Figure 25 ). P3 endopod ( Fig. 19F, G View Figure 19 ) distinctly three-segmented; enp-1 small, without ornamentation or armature; enp-2 short, with serrate, posterior seta and rigid, sigmoid apophysis arising from anterior surface; distal third of apophysis with denticulate margin; enp-3 a short bulbous segment, with one short, pinnate seta apically.
Fifth legs ( Fig. 18D View Figure 18 ) with baseoendopod and exopod fused into a common elongate plate, tapering distally towards a long, bare, spinous process, which is longer than the plate; inner margin with serrate spine; outer margin with small serrate spine, three naked setae and sparsely plumose seta derived from baseoendopod; anterior surface with three large tube-pores.
Sixth pair of legs ( Figs 18E View Figure 18 , 25C View Figure 25 ) asymmetrical, with two long, sparsely pinnate setae and one short, bare seta each. Left or right leg articulating according to sinistral or dextral development of testis and vas deferens. First postgenital somite with pattern of transverse spinular rows near ventral anterior margin ( Figs 18E View Figure 18 , 25C View Figure 25 ).
Caudal ramus ( Fig. 21F View Figure 21 ) as in female.
Remarks: Mielke (1975) expressed some reservations about the identity of his Evansula material from the Isle of Sylt. He decided to attribute his specimens to E. pygmaea on the following grounds: (1) body length shorter than in E. incerta , (2) distal spines of P1 enp- 2 claw-like, and (3) similarity in female P5 with Scott’s (1903a) original description. A comparison based on one female and one male from Sylt revealed no differences with the type material of E. cumbraensis except for (1) the P2 endopod, which, as well as its distal spine, appears to be slightly longer in the German specimens ( Fig. 20C View Figure 20 ), (2) the shape of the geniculate spines on P1 enp-2, which appear to be slightly more stout ( Fig. 19C View Figure 19 ). However, the precise length and shape of these elements are dependent on their orientation on the slide (cf. Fig. 19A, B View Figure 19 ), and (3) small differences in caudal ramus contours and length of styliform part of seta V (but see under variability). Mielke seems to have found only E. cumbraensis . However, other studies reporting both E. pygmaea and E. incerta ( Noodt, 1952, 1957) potentially indicate the occurrence of other sympatric species on the Isle of Sylt.
Martínez Arbizu & Moura’s (1994) drawings of the genital field (cf. position and size of copulatory pore; however, the innermost setule on P6 was overlooked) and the P2 of both sexes (length of exopodal spines, endopod in male) of E. pygmaea from Sylt leave no doubt that they were also dealing with E. cumbraensis .
Mielke (1975) commented on the structure of the ‘plumose’ setae found on the female P5. He described the pinnules as composed of ‘... einem kurzen, verstärkten Basalteil und einem langen Endteil, der leicht abbrechen kan. Bei Verlust einer grösseren Anzahl von Fiederendstücken werde kurz behaarte Borsten vorgetäuscht’. Scanning electron microscopy revealed that these plumose setae are clearly pinnate ( Fig. 26D View Figure 26 ) and the plumosity is an artefact created by epibiotic filamentous bacteria attached at and around the short pinnules ( Fig. 26C View Figure 26 : distal part of second innermost seta). McAllen & Hannah (1999) observed similar biofouling by filamentous bacteria on the exoskeleton of the high-shore rockpool harpacticoid Tigriopus brevicornis (O.F. Müller) . In this species, the major biofoulant is Leucothrix mucor OErsted, which tends to colonize between the body tergites where the exoskeleton is at its thinnest for the action of chitinolytic enzymes. The filamentous bacteria found in E. pygmaea , E. cumbraensis , and E. arenicola appear to show a high site specificity, attaching themselves only to the long setae of the P5 (usually the female).
Variability: Found only in the proximal styliform part of seta V, the length and shape of which can vary considerably within a population. Figure 21G View Figure 21 illustrates the intraspecific variation found in the Elie population. The Sylt population clearly fits within this range ( Fig. 21E View Figure 21 ; Mielke, 1975: abb. 66B).
Differential diagnosis: The most important diagnostic features of female E. cumbraensis are the elongated exopodal spines on the middle and distal segments of P2 and the very large copulatory pore. It shares with E. incerta the absence of ventral spinular rows near the anterior margin of the anal somite in the female. Males of the closely related E. cumbraensis and E. polaris can be distinguished by the detailed morphology of the P3 endopod (segmentation, shape of enp-3) and P5 (spinous process smooth vs. pinnate).
Distribution: Scotland: Fintray Bay, Isle of Cumbrae (present account); Elie in Firth of Forth (present account).
Germany: Isle of Sylt ( Mielke, 1975, 1976; Martínez Arbizu & Moura, 1994).




| NHM |
University of Nottingham |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |