
Evansula pygmaea
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2006.00227.x |
|
persistent identifier |
https://treatment.plazi.org/id/03F487A0-FFA3-FFB6-FFD3-F9A7FDC99052 |
|
treatment provided by |
Felipe |
|
scientific name |
Evansula pygmaea |
| status |
|
EVANSULA PYGMAEA (T. SCOTT, 1903A)
Synonyms: Tetragoniceps pygmaeus T. Scott, 1903a ; Evansia pygmaea (T. Scott, 1903a) Scott (1906a) ; Evansula pygmaea (T. Scott, 1903a) Scott (1906b) .
Original description: Scott (1903a: 117–118, plate IV, figs 11–19).
Additional descriptions: None. The additional illustrations given by Mielke (1975) and Martínez Arbizu & Moura (1994) are based on specimens of E. cumbraensis sp. nov.
Type locality: Scotland, Firth of Forth , near Musselburgh .
Material examined: NHM, reg. nos 45033–035: Firth of Forth , Scotland; 1 ♀ in alcohol (labelled ‘cotype’); coll. T. Scott, 1894; as part of Cannon A. M. Norman collection (1911.11.8); dissected on eight slides .
Redescription
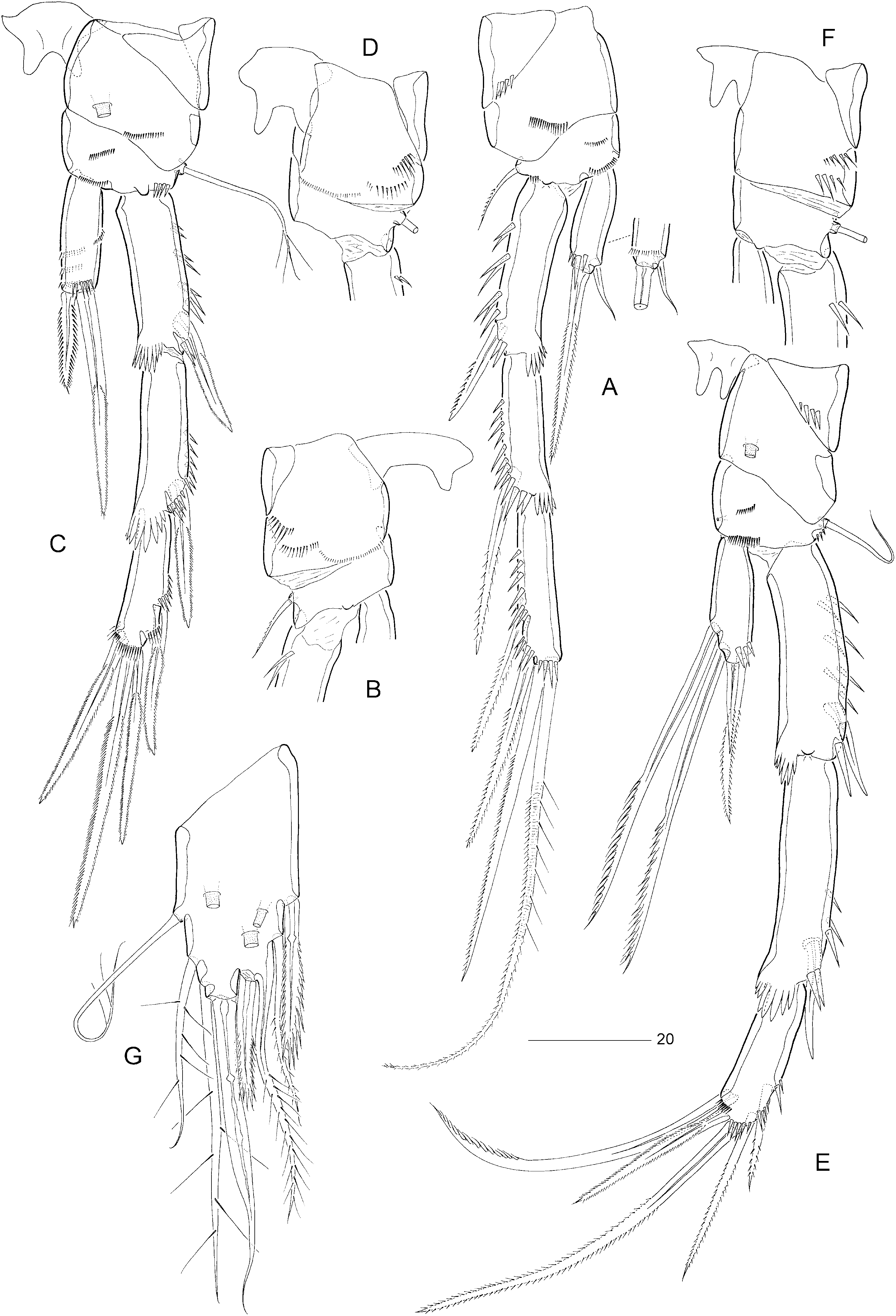
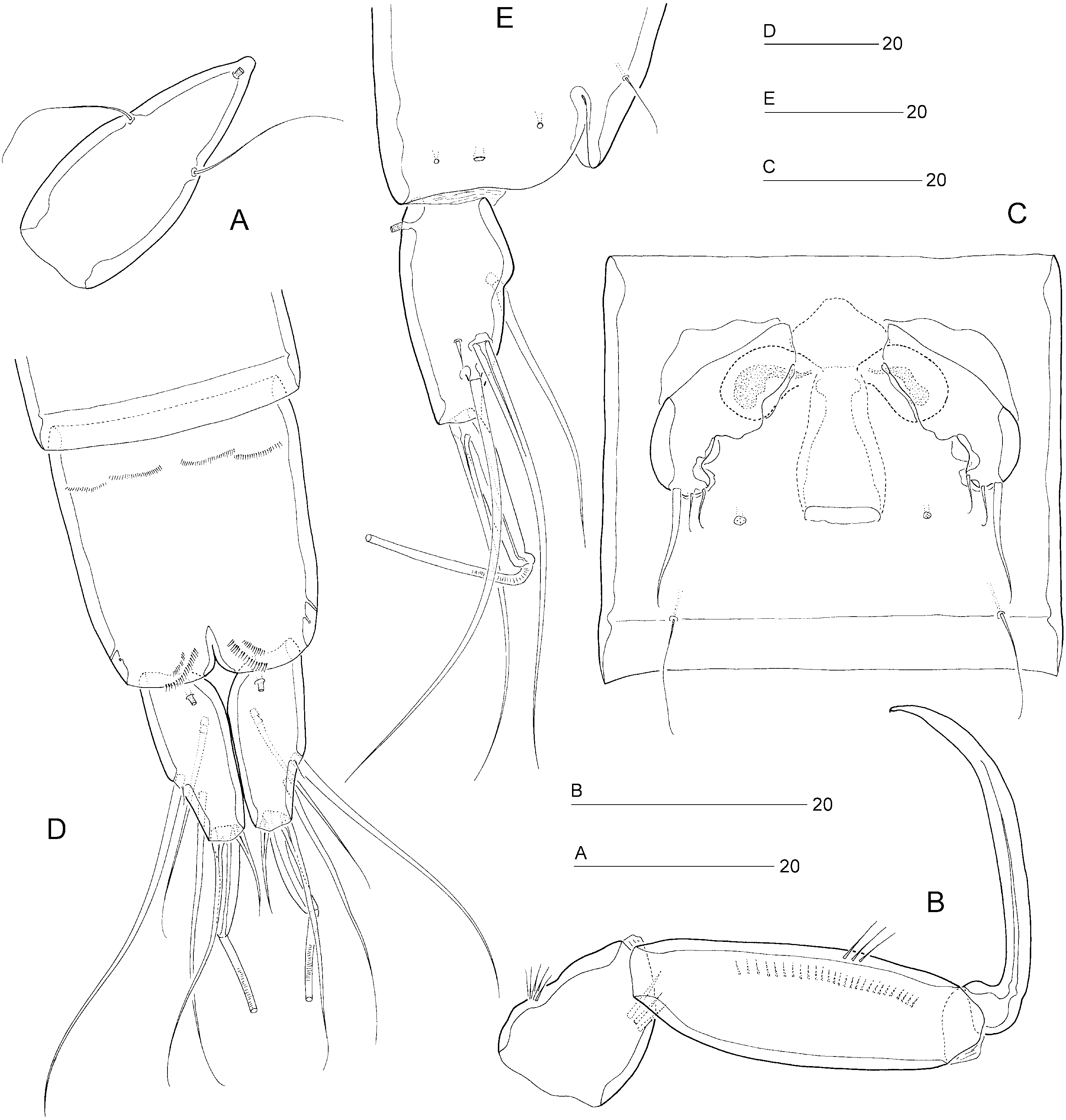
Female: Total body length: 475 µm. Body slender, cylindrical ( Fig. 9A, B View Figure 9 ), semitransparent, yellowish; no distinct separation between prosome and urosome. Genital double-somite completely fused ( Figs 9A, B View Figure 9 , 12C View Figure 12 ); original segmentation marked dorsally by paired anterior and posterior sensillae ( Fig. 9A, B View Figure 9 ). Anal somite only slightly longer than wide (50 × 45 µm), with three pairs of secretory pores laterally ( Fig. 12D, E View Figure 12 ); ventral surface with four rows of tiny spinules near anterior border ( Fig. 12D View Figure 12 ); posterior margin with two short spinular rows on either side of ventral midline ( Fig. 12D View Figure 12 ). Anal operculum weakly developed, unarmed ( Figs 10E View Figure 10 , 12E View Figure 12 ).
Caudal rami slightly convergent ( Figs 9A View Figure 9 , 10E View Figure 10 , 12E View Figure 12 ); distinctly constricted around anterior rim resulting in strongly convex contour of proximal inner margin; length (measured along outer margin) approximately 2.8 times proximal width; dorsal surface without chitinous spur but with raised spinular row anterior to seta VII ( Fig. 10E View Figure 10 ) and with blunt dorsal process (discernible in lateral aspect: Fig. 12E View Figure 12 ); with seven setae, setae I–VI in distal and seta VII in proximal half ( Fig. 10D, E View Figure 10 ); seta I diminutive; setae II–III long and bare; seta IV long and naked, more than twice the length of styliform part of V; seta V long, with distinct flexure zone between styliform part and long distal flagellate part, fused at base with seta IV; seta VI vestigial and spiniform; seta VII triarticulate at base, somewhat flattened near tip ( Fig. 10E View Figure 10 ) and located along proximal inner margin; ventral and dorsal surfaces with one tube-pore each ( Fig. 12D, E View Figure 12 ).
Rostrum elongate ( Fig. 12A View Figure 12 ), with slightly convex margins in proximal half, tapering distally; distinctly shorter than first antennulary segment ( Fig. 9A View Figure 9 ); with two long sensillae; median pore positioned dorsally near apex of rostrum.
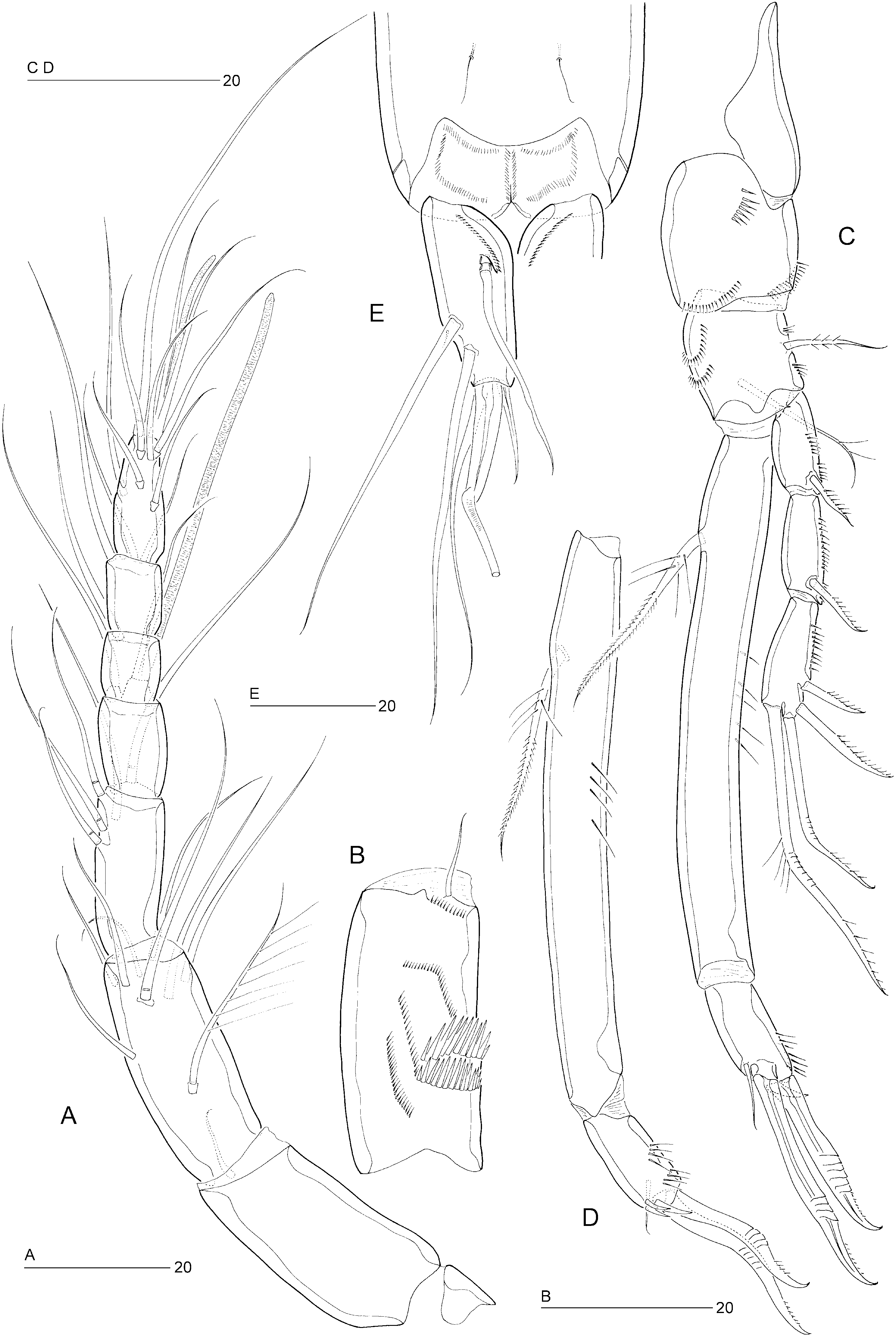
Antennule seven-segmented ( Fig. 10A View Figure 10 ). Segment 1 only slightly shorter than segment 2, with small sclerite around proximal posterior margin, anterior surface with pattern of spinular rows, as illustrated in Figure 10B View Figure 10 ; segment 2 longest, without secretory pore; segment 4 with distal cylindrical process bearing large aesthetasc (55 µm). Armature formula: 1-[1], 2-[8 + 1 pinnate], 3-[5], 4-[1 + (1 + ae)], 5-[1], 6-[3], 7-[7 + acrothek]. Apical acrothek consisting of two long setae and one slender aesthetasc (25 µm).
Antenna, mandible, maxillule, and maxilla as in E. incerta .
Maxilliped ( Fig. 12B View Figure 12 ) well developed, subchelate, directed inwards. Syncoxa well developed, with two spinular rows, without seta. Bases elongate, with two to three long spinules on anterior surface and a spinular row along posterior inner margin. Endopod represented by strong, curved, bare claw.
P1 ( Fig. 10C, D View Figure 10 ). Praecoxa strongly developed, unarmed. Coxa with three spinular rows. Basis with long, semiplumose, inner seta and short, pinnate, outer seta. Exopod three-segmented; with two spines and two geniculate setae on exp-3. P1 endopod prehensile, distinctly longer than exopod; proximal segment approximately nine times as long as average width, with pinnate inner seta being plumose in proximal third; distal segment short, with subdistal setule, and two geniculate spines distally.
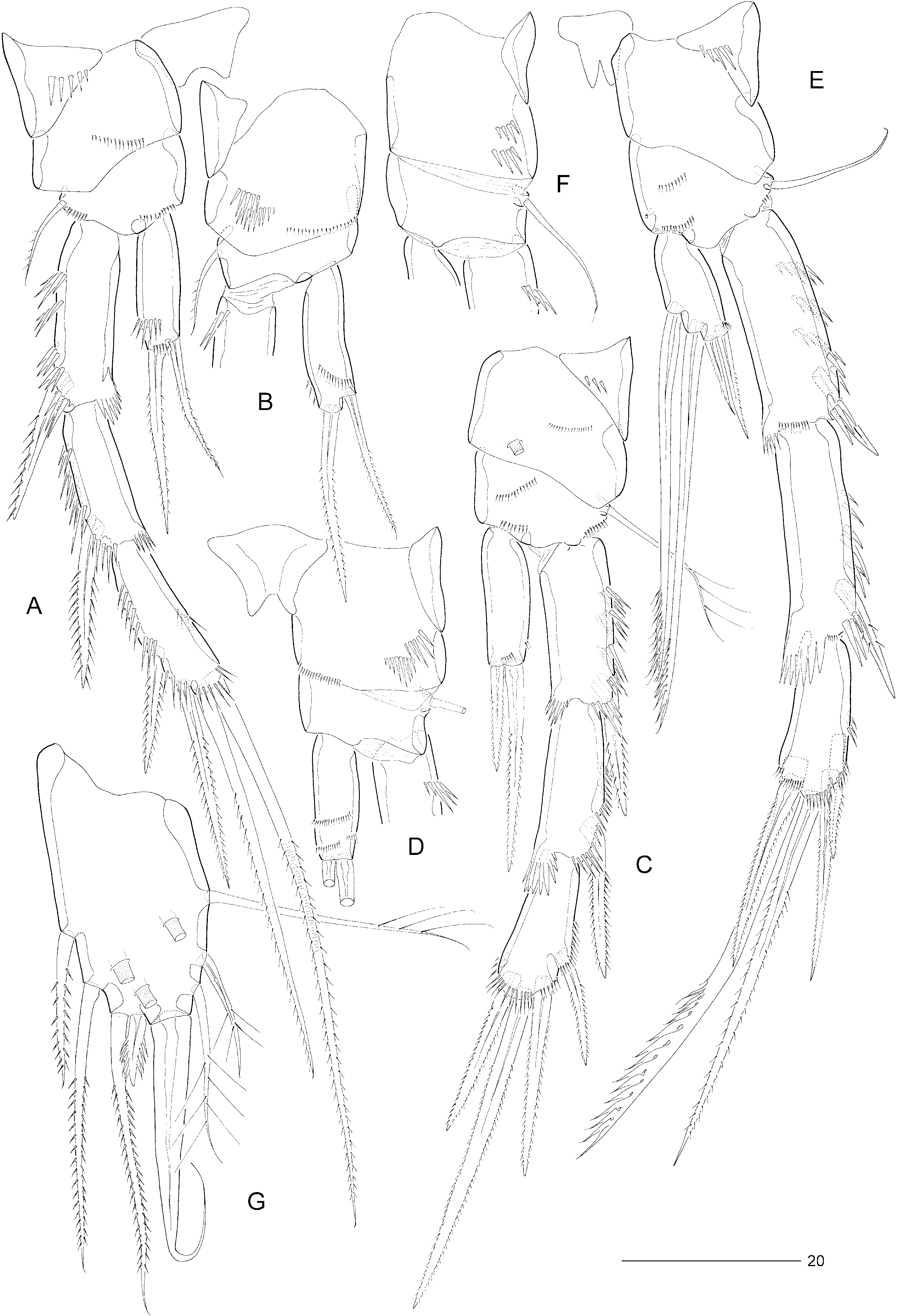
Swimming legs P2–P4 ( Fig. 11A–F View Figure 11 ) somewhat more robust than in E. incerta . Praecoxae well developed, with spinular row on anterior surface ( Fig. 11A, C, E View Figure 11 ). Coxae with pattern of spinules as in Figure 5A–F View Figure 5 , with large tube-pore on anterior surface of P3 (but not P4). Bases with outer seta (short and pinnate in P2, long and plumose in P3, long and bare in P4); with spinular rows on anterior surface only ( Fig. 11A, C, E View Figure 11 ). Exopods three-segmented, endopods one-segmented. Exopodal spines of P3 pinnate ( Fig. 11C View Figure 11 ). P2 endopod approximately 0.8 times as long as exp-1; with two spinular rows on anterior and one spinular row on posterior surface; inner element setiform, pinnate, only slightly shorter than distal spine. Armature elements of P3 endopod spiniform; inner spine approximately half length of outer spine. Seta and spine formulae as for genus.
Fifth pair of legs ( Fig. 11G View Figure 11 ) with baseoendopod and exopod fused into a common elongate plate, tapering distally; apex with strong, articulating spine, slightly longer than plate and with flagellate tip; outer margin with three sparsely plumose setae (including seta derived from baseoendopod), middle and distal ones distinctly shorter than in E. incerta ; inner margin with three pinnate setae (middle one longer than in E. incerta ) and one short serrate spine (much shorter than in E. incerta ); none of armature elements fused to plate; anterior surface with three large tube-pores. Inner margin setae sometimes with foreign tubular filaments attached to individual pinnules.
Sixth legs ( Fig. 12C View Figure 12 ) each represented by a small operculum closing off gonopore; armature consisting of setiform outer element and two accessory setules. Genital apertures not fused medially; copulatory pore large, located slightly posterior to gonopores; leading via short chitinized copulatory duct to paired anterior extensions, positioned anterior to genital apertures; copulatory pore flanked by two small secretory pores.
Male: Based on Scott’s (1903a) original description.
P3 endopod distinctly three-segmented; enp-1 small and unarmed; enp-2 drawn out into long, bent apophysis (the accessory seta on this segment was probably overlooked by T. Scott); enp-3 with one short apical seta.
P5 baseoendopod and exopod fused into common triangular plate, tapering distally towards curved, naked, spinous process, which is slightly longer than the plate; inner margin with one seta; outer margin with five armature elements, the distalmost one short-
est and setiform; no details given about the ornamentation of these elements.
Remark: The plumosity of the long inner setae on the female P 5 in Scott’s (1903a) original description is an artefact, being caused by attached filamentous bacterial epibionts.
Differential diagnosis: Perhaps the most characteristic feature of E. pygmaea is the peculiar shape of the slightly convergent caudal rami with their strongly convex proximal inner margins. The species is most closely related to E. cumbraensis sp. nov. and E. polaris sp. nov. with which it shares the raised spinular row on the caudal ramus. E. pygmaea differs from E. cumbraensis sp. nov. in the length of the outer spines on P2 exp-2 and -3, the size of the copulatory pore, the presence of an anteroventral spinular row on the female anal somite and in both size and ornamentation of the setae on the female P5 (in particular the short inner distal spine). The major discrepancy with the Arctic species is found in the shape and length of the caudal ramus. Provided Scott’s (1903a) illustration (plate IV, Fig. 17 View Figure 17 ) is accurate, additional differences can be found in the male P5.
Distribution: Scotland: Musselburgh in Firth of Forth ( Scott, 1903a, 1906b).
The following list contains all records attributed to E. pygmaea that do not provide enough information for a positive identification to be made.
Scotland: Firemore, Loch Ewe ( McIntyre & Murison, 1973), River Ythan ( Hockin, 1982c).
Wales: Trearddur Bay, Traeth Lligwy, Traeth Bychan, and Tal-y-Foel, Anglesey ( Geddes, 1972).
England: Whitsand Bay, Cornwall ( Harris, 1972a, b, c), St. Andrews and St. Agnes, Isles of Scilly ( Wells, 1961, 1970), River Exe ( Wells, 1963).
Ireland: Silver Strand, Galway ( Bodin & Jackson, 1989; Holmes & O’Connor, 1990).
Germany: Isle of Sylt ( Noodt, 1952, 1957), Kieler Bucht ( Klie, 1929, 1950; Remane, 1933; Kunz, 1935; Scheibel, 1972, 1973; Scheibel & Noodt, 1975; Anger & Scheibel, 1976), Bottsand and Weissenhaus in Kieler Bucht ( Noodt, 1956, 1957), Bocknis Eck in Kieler Bucht ( Scheibel, 1976), Helgoland ( Kunz, 1938; Klie, 1950), Elbe estuary ( Riemann, 1966).
The Netherlands: Oosterschelde ( Smol et al., 1994), Westerschelde estuary ( Van Damme, Heip & Willems, 1984).
Belgium: Sluice Dock, Ostend ( Thielemans & Heip, 1984), Kwinte Bank ( Willems et al., 1982a, b), coastal zone ( Heip, Herman & Vincx, 1984).


| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |