
Eunice taoi, Hsueh, Pan-Wen & Li, Yan-Huei, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3802.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:20400951-A62C-49FD-9D70-3FE54557E5D8 |
|
DOI |
https://doi.org/10.5281/zenodo.6141568 |
|
persistent identifier |
https://treatment.plazi.org/id/AF67D207-FFCC-FFBF-FF14-FBD5FC2AA74E |
|
treatment provided by |
Plazi |
|
scientific name |
Eunice taoi |
| status |
sp. nov. |
Eunice taoi View in CoL sp. nov.
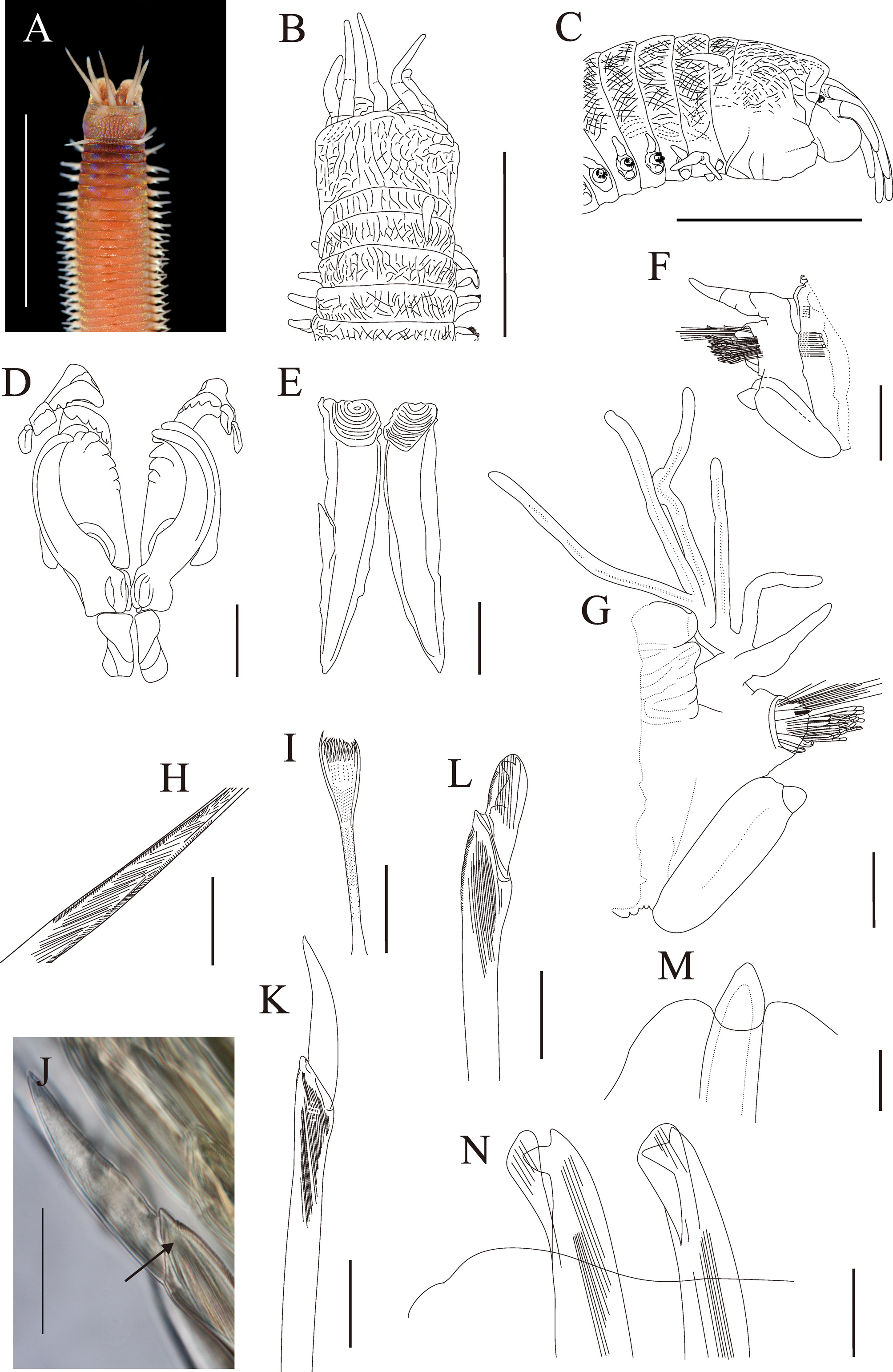
Fig. 8 View FIGURE 8
Material examined. Holotype ( NMNS 6896-001), Jihuei (23°06´53˝N, 121°24´11˝E), Taitung County, intertidal algal and coral mixed reef, August 23, 2010; paratypes: three specimens ( NMNS 6896-005, 006~007), Jihuei (23°06´53˝N, 121°24´11˝E), Taitung County, intertidal algal and coral mixed reef, October 18, 2009 and August 22, 2010, respectively; three specimens ( NMNS 6896-012~013; AM W45884), Wanlitong (21°59´46˝N, 120°42´12˝E), Pingtung County, intertidal coral reef, November 13, 2009; three specimens ( NMNS 6896- 0014~016), Jialulan, Taitung County, rocky intertidal, August 11–12, 2010; one specimen ( NMNS 6896-017), Shanyuan (22°50´19˝N, 121°11´19˝E), Taitung County, intertidal algal reef, August 23, 2010; two specimens ( NMNS 6896-018; AM W45885), Shitiping (23°28´59˝N, 121°30´47˝E), Hualien County, rocky intertidal, August 27, 2010; seven specimens ( NMNS 6896-002~004, 008~011), Jihuei (23°06´53˝N, 121°24´11˝E), Taitung County, intertidal algal and coral mixed reef, October 14, 2007 and August 23, 2010, respectively.
Description. Holotype, female with egg, complete specimen but broken in three parts, 561 mm in length for 633 chaetigers; maximum width 4.1 mm at chaetiger 53; length through chaetiger 10 about 9.7 mm, width at chaetiger 10 about 3.6 mm; one pair anal cirri, slender tapering, without articulations.
Prostomium distinctly shorter than peristomium, equal width and about 1/2 as deep as peristomium, prostomial lobes frontally rounded, dorsally flattened, median sulcus deep ( Fig. 8 View FIGURE 8 A–C). Eyes present, situated posteriorly to base of palps, ovate and greyish purple. Palps and antennae thick, arranged in a semicircle, median antennae separated from lateral antennae; palpophores and ceratophores base short cylindrical, wide than long, with one indistinct ring-shaped articulation; palpostyles and ceratostyles slender tapering, surface with numerous papillae, without articulations; A-I, A-II and A-III similar length, palps distinctly shorter than antennae; A-II reaching to chaetiger 2, A-I and A-III to chaetiger 1, palps to posterior margin of anterior peristomial ring. Peristomium inflated anteriorly, lower lip muscular; separation between rings distinct dorsally and ventrally; anterior ring about 3/4 of total peristomial length; peristomial cirri tapering, without articulations, reaching middle of anterior peristomial ring ( Fig. 8 View FIGURE 8 A–C).
Maxillary formula: 1+1, 6+5, 6+0, 3+8, 1+1; Maxillary teeth short, MxIII located at front-end part of distal arc with left MxIV ( Fig. 8 View FIGURE 8 D); mandibles flat ( Fig. 8 View FIGURE 8 E).
Branchiae present, pectinate ( Fig. 8 View FIGURE 8 G), present from chaetiger 77 to chaetiger 630, representing more than 65% of chaetigers branchiate; 1st to 6th and the last pairs of branchiae with single filament, maximum 6 filaments present from about chaetiger 133–304, not reduced in mid-body region; branchial stems erect, flexible, filaments slender, distinctly longer than dorsal cirri ( Fig. 8 View FIGURE 8 G).
Anterior neuropodial acicular lobes bilobed, upper lobes distally rounded, lower lobes obliquely truncate ( Fig. 8 View FIGURE 8 F), median and posterior acicular lobes not bilobed, triangular aciculae emerging above midline ( Fig. 8 View FIGURE 8 M); prechaetal lobes small, transverse folds; anterior postchaetal lobes small, folds following outline of acicular lobes, posterior postchaetal lobes small, transverse folds at middle and posterior segments. First five ventral cirri digitiform, subsequently basally strongly inflated to about chaetiger 133, inflated bases thick, transverse welts, narrow tips short and button-shaped; inflated bases becoming ovate to spherical by chaetiger 134, basal inflation gradually reduced on subsequent chaetigers, becoming slender and digitiform on posterior chaetigers. Anterior dorsal cirri tapering, posterior dorsal cirri slender tapering, all without articulations ( Fig. 8 View FIGURE 8 F–G).
Limbate chaetae elongate, longer than all other chaetae, narrow, marginally serrated, internal striations present ( Fig. 8 View FIGURE 8 H). Pectinate chaetae furled, flaring; shafts slender, surface pilose, one marginal teeth heterodont, 12–14 inner teeth ( Fig. 8 View FIGURE 8 I). Compound spinigers present from chaetiger 1 to chaetiger 83; shafts slightly inflated, marginally serrated, internal striations present ( Fig. 8 View FIGURE 8 K); appendages thick, knife-shaped, proximal nearly parallel sides, distal slightly curved, length/width ratio (n=4) about 4.06. Compound falcigers replacing compound spinigers from about chaetiger 83; shafts distally inflated, marginally serrated ( Fig. 8 View FIGURE 8 L); appendages small, short, length/width ratio (n=4) is about 3.83, with nearly parallel sides, tapering and distinct heads, bidentate; proximal tooth larger than distal tooth, triangular, directed laterally; distal tooth gently curved; guards almost symmetrically rounded, marginal serrations and internal striations present, without mucros ( Fig. 8 View FIGURE 8 L); pseudo-compound falcigers absent. Aciculae light brown to black on anterior segments, amber on posterior segments, thick and straight, tapering with blunt tip, cross-sections rounded, not mucronate ( Fig. 8 View FIGURE 8 M), paired aciculae present from chaetiger 1 to 45, on subsequent chaetigers only one present; core and sheath distinctly separated in aciculae and subacicular hooks. Subacicular hooks yellow, bidentate, about as thick as aciculae ( Fig. 8 View FIGURE 8 N), present from chaetiger 71 to last chaetiger, single in chaetigers 71 to 78 and 261 to 633, paired in chaetigers 79 to 260; shafts of hooks straight, distally tapering; proximal tooth larger than distal tooth, triangle, directed laterally; distal tooth directed obliquely; guards covering only proximal tooth or both teeth ( Fig. 8 View FIGURE 8 N).
Etymology. The name is derived from the Tao aboriginal tribe on Orchid Island, Taiwan, as a tribute to their oceanic cultures.
Type locality. Jihuei, Taitung County, Taiwan
Habitat. The worm appears to capable of making a burrow in coral or coral and algal mixed reefs and building leathery tube which protrude a few to several tens of centimeters above the reefs. However, some individuals utilize burrows of the boring bivalve, Jouannetia globulosa (Quoy & Gaimard, 1835) (Family Pholadidae ), to build tube in sandstone and shale.
Distribution. Known only from Taiwan, occurs in low intertidal and subtidal habitats on eastern and southern coasts of Taiwan.
Colour. In life, anterior body of the worm is orange red with purplish iridescence on the dorsum ( Fig. 8 View FIGURE 8 A). This orange red colour fades away at the middle portion of the body and is replaced by whitish with light orange on both sides of the posterior body. Eyes are dark brown colour.
Remarks. Eunicid polychaetes with compound spinigers are often referred to the genus Euniphysa Wesenberg-Lund, 1949 ( Fauchald 1977: 106) . However, Fauchald (1992) noted that three species in the genus Eunice Cuvier, 1817 , namely Eunice impexa Grube, 1878 , Eunice tubicola (Treadwell, 1922) and Eunice tubifex Crossland, 1904 , also have compound spinigers, although E. tubifex was transferred to the genus Euniphysa by Miura (1986). In the study of phylogeny and biogeography of the genus Euniphysa Wesenberg-Lund, 1949 , Lu & Fauchald (2000) argued that some Eunice spp. may have both compound spinigers and falcigers as in members of Euniphysa , but those Eunice spp. differed from members of Euniphysa in having short, triangular maxillae teeth (vs. long pointed maxillae teeth), short and distinctly articulated prostomial antennae (vs. slender and smooth prostomial antennae). On the basis of the above arguments, they proposed a new combination of Euniphysa misakiensis Miura, 1987 as Eunice misakiensis ( Miura 1987) and acknowledged four species ( E. impexa , E. misakiensis , E. tubicola and E. tubifex ), which have compound spinigers as belonging to this genus. The present study adds another species to this group of Eunice .
Of above-mentioned four Eunice species, Eunice tubifex resembles E. taoi sp. nov. Both species have nonarticulated ceratostyles, deep medial sulcus and pectinate branchiae ( Fauchald 1992: 327, Fig. 112a, d, i, n; Fig. 8 View FIGURE 8 B, G). However, they are morphologically different in terms of the form of ceratostyle of prostomial appendages, peristomial cirri, pectinate chaetae and compound spiniger appendages. Eunice tubifex have digitiform ceratostyles, thick and basally inflated peristomial cirri, furled and tapering pectinate chaetae and more slender compound spiniger appendages ( Fauchald 1992: 327, Fig. 112a, b, f, i–j, l, r), in comparison to smooth and tapering ceratostyles, tapering peristomial cirri, furled and flaring pectinate chaetae and much shorter and robust compound spiniger blades of E. taoi sp. nov. ( Fig. 8 View FIGURE 8 B–C, I, K). Eunice misakiensis is another species that also resembles E. taoi sp. nov. However, it can be separated from E. taoi sp. nov., by the shape of the median sulcus, the pectinate chaetae and by the compound spiniger appendages. The former species possesses an incised median sulcus, flat and flaring pectinate chaetae and far more slender compound spiniger appendages ( Miura 1987: 6, Fig. 4 View FIGURE 4 a–b, i, k, l; Imajima 2007: 353, Fig. 108a, j, l), in contrast to the deep median sulcus, furled and flaring pectinate chaetae and much shorter and robust compound spiniger appendages of E. taoi sp. nov. ( Fig. 8 View FIGURE 8 B, I, K). Lastly, E. taoi sp. nov., have uniquely short and robust compound spinigers that is not seen in the other four species of this group. The length/width ratios of compound spinigers are 7.7, 11.3 or 14.4, 13.0, 9.6 or 11.0 and 5.8 (n=10) for E. impexa , E. misakiensis , E. tubicola , E. tubifex and E. taoi sp. nov., respectively (measurements for the first four species were from Miura 1987, Fig. 4 View FIGURE 4 l; Fauchald 1992, Fig. 57c, 111c, 112b, j; Imajima 2007, Fig. 108j).
Eunice taoi sp. nov., is the largest worm among the Eunice species discussed in the present study, with some individuals reaching over 710 mm. The body size and the length of the worm tube of Eunice taoi sp. nov., appears to vary within the different habitats which it occupies. Worms collected from sandstone and shale habitats tend to be much smaller in body size and shorter in tube length than those from coral and algal mixed reefs. Presumably, the body size and tube length of the worm are dependent on the depth of burrows made by J. globulosa , which is often only a few centimeters deep. This may suggest that Eunice taoi sp. nov. cannot bore into shale rocks but can utilize burrows made by the bivalve. However, they appear capable of boring deep into coral reefs; in many collection attempts, we traced those leathery worm tubes deep inside coral reefs by digging out the entire worm tube. However, those worms are still out of reached and hiding deep in tubular burrows. Some of larger sized worms were collected at depths of more than 30 cm deep within the reefs.
| NMNS |
National Museum of Natural Science |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.