
Echiniscus spinulosus ( Doyère, 1840 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5344.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:DCF48473-AC31-4CDB-808F-453F8F280002 |
|
DOI |
https://doi.org/10.5281/zenodo.8349272 |
|
persistent identifier |
https://treatment.plazi.org/id/8119D633-B96F-FFC1-1CED-F937BC1CFCEC |
|
treatment provided by |
Plazi |
|
scientific name |
Echiniscus spinulosus ( Doyère, 1840 ) |
| status |
|
18. Echiniscus spinulosus ( Doyère, 1840) View in CoL
Figures 30–33 View FIGURE 30 View FIGURE 31 View FIGURE 32 View FIGURE 33 , 40 View FIGURE 40 , Tables 8–9 View TABLE 8 , Supplementary Material 9
Synonyms
Echiniscus carusoi Pilato, 1972 View in CoL syn. nov.
Emydium spinulosum Doyère, 1840 View in CoL
Locus typicus: France, Saint-Maur-des-Fossés.
New type locality: 38°7’23”N, 2°34’51”W, 1459 m asl: Spain, Andalucía, Sierras de Cazorla, Segura y Las Villas. Moss from rock in a pine forest. Found together with E. testudo .
The neotype series consists of 33 specimens: 23 ♀♀ and 10 juveniles mounted on permanent slides ES.226.01– 8 (neotype: ES.226.06). 8 specimens used for DNA extraction; 50 specimens on SEM stub 22.07.
Additional localities: See Table 1 View TABLE 1 for all sequenced populations. Verified and trustworthy records: (1) Switzerland ( Rahm 1928); (2) Italy, Piedmont, Val di Susa, Avigliana ( Bertolani 1941); (3) Italy, Veneto, Belluno, vicinity of Lago di Vedana ( Ramazzotti 1945); (4) Romania: Suceava County, the Rarău Massif, the Giumalău Massif, Inău, Neamț County, the Ceahlău Massif, Hunedoara County, the Retezat Massif, and Prahova County, Sinaia ( Rudescu 1964); (5) Austria, East Tirol ( Mihelčič 1969); (6) Italy, Veneto, Basalovo (all records of E. spiniger from Basalovo belong to E. spinulosus ; Durante & Maucci 1972); (7) Slovenia, Gračišče (misidentified E. spiniger ; Maucci 1972); (8) Italy, Friuli-Venezia Giulia, Carso Triestino, Sales, Basovizza, Monte Grisa, Monte San Leonardo, Monte Spaccato, Padriciano, Prosecco (all records of E. spiniger from Carso Triestino belong to E. spinulosus ; Maucci 1973–4); (9) France, Cévennes, Mont Aigoual ( Bertrand 1975); (10) Great Britain: England, Warwickshire, Northend, Wales, Pembrokeshire, Broad Haven, and Scotland, Oban (summary in Morgan & King 1976); (11) Greece, Delphi, Megalopolis ( Durante Pasa & Maucci 1979a); (12) France, Nice ( Séméria 1982); (13) Greece, Parnassós (misidentified E. spiniger ; Maucci & Durante Pasa 1982); (14) Italy, Sardinia, Oliena ( Binda & Guglielmino 1982); (15) Spain, Andalucía, El Zegri and Atajate (Maucci & Durante Pasa 1984); (16) Portugal, vicinity of Coimbra, Santa Luzia ( Maucci & Durante Pasa 1985); (17) Slovenia, vicinity of Lake Cerknica ( Čuček 1985); (18) Italy, Puglia, San Marco in Lamis ( Maucci 1986); (19) Austria, Steiermark ( Dastych 2015); (20) ca. 48°24’N, 2°42’E, 70 m asl: France, Fontainebleau (6 ♀♀; an examined population nearest to the type locality; Gąsiorek & Degma 2018).
New records:
• ca. 45°39’48’’N, 13°59’21’’E, 412 m asl: Slovenia, Škocjanske jame, coll. on 20 th May 1972 (the Maucci collection, not reported in literature and misidentified as E. spiniger );
• ES.133; 40°13'59"N, 2°3'46"W, 1102 m asl: Spain, Castilla-La Mancha, Parque Natural Serrania de Cuenca, mosses from rocks in maritime pine ( Pinus pinaster ) forest, Piotr Gąsiorek & Witold Morek coll. on 16 th April 2018 (1 ♀);
• ES.157; 37°50'48"N, 5°15'44"W, 286 m asl: Spain, Andalucía, Sierra de Hornachuelos, Las Herrerías, moss from rock in maquis shrubland, Piotr Gąsiorek & Witold Morek coll. on 18 th April 2018 (2 ♀♀);
• FR.151; France, Lozère, Massegros-Causses-Gorges, Point Sublime, moss from a concrete wall in maquis shrubland, Thomas Pape coll. on 1 st August 2022 (1 ♀).
Etymology: From Latin spīnulōsus = spiny, referring to the presence of numerous spiniform trunk appendages. An adjective in nominative singular.
Redescription. Females (i.e. from the third instar onwards; measurements and statistics in Table 8 View TABLE 8 ). Medium-sized to large (ca. 240–320 µm in length, but can reach even up to 350 µm in other populations), compact body ( Figs 30–31 View FIGURE 30 View FIGURE 31 ), orange to red and with large red eyes, dissolving in Hoyer’s medium. Dactyloid cephalic papillae (secondary clavae) and (primary) clavae; cirri growing out from bulbous cirrophores ( Figs 30 View FIGURE 30 , 31B–C View FIGURE 31 ). Cirri A relatively long (ca. 50% of the body length on average). Body appendage configuration A-(B)- C-Cd- D-Dd- E; appendages Cd and Dd developed in a form of long spines, several times longer than minute, often triangular lateral spicules ( Figs 30–31 View FIGURE 30 View FIGURE 31 ). Asymmetries in the development of trunk appendages frequent ( Fig. 30A View FIGURE 30 ).
Dorsal plates with the spinulosus sculpturing: large pores dominating the entire dorsal armour ( Figs 30–31 View FIGURE 30 View FIGURE 31 ). Cephalic plate with an anterior chalice-like incision ( Fig. 30 View FIGURE 30 ); cervical plate narrow, but sculptured and clearly discernible from scapular plate ( Figs 30 View FIGURE 30 , 31B–C View FIGURE 31 , 32A View FIGURE 32 , 33A View FIGURE 33 ). Scapular plate broad, without a median suture, but with two lateralmost sutures/ridges that delineate rectangular portions at the level of cirri A ( Figs 31B–C View FIGURE 31 , 32A View FIGURE 32 ). Median plates 1 and 3 unipartite, the latter greatly reduced and partially hidden under overlapping posterior portions of pII and anterior portion of caudal plate; m2 bipartite, but the anterior portion is poorly developed and partially hidden under overlapping posterior portions of pI. Paired segmental plates I–II with well-delineated anterior and posterior parts by narrow and deep transverse belts ( Figs 30–31 View FIGURE 30 View FIGURE 31 , 32B View FIGURE 32 ). Anterior portions of pI–II and m2 with reduced endocuticular matrix, so spaces between pores are thinner and, consequently, pores are divergent phenotypically compared with other plates ( Figs 30–31 View FIGURE 30 View FIGURE 31 , 32B View FIGURE 32 ). Caudal plate with two long indistinct incisions, epicuticular ridges absent ( Figs 30–31B View FIGURE 30 View FIGURE 31 , 32C View FIGURE 32 ).
Venter uniformly granulated; subcephalic and genital plates in the form of dark trapezoidal/oval shapes may be present. Hexapartite gonopore placed between genital plates, and trilobed anus between legs IV. Legs massive, with large and sculptured (pores identical with dorsal ones) pedal plates in median portions ( Figs 31B–C View FIGURE 31 , 33E–F View FIGURE 33 ) and pulvini in proximal portions ( Figs 31B–C View FIGURE 31 ). Spine on leg I ( Figs 31B–C View FIGURE 31 , 33B, E View FIGURE 33 ) and papilla on leg IV present ( Figs 31B–C View FIGURE 31 ). Claws large and heteronych ( Figs 33B–F View FIGURE 33 ): claws I–III of equal height, but claws IV considerably higher. External claws on all legs smooth; internal claws I–III with primary spurs of a typical size and positioned at ca. 20% of the claw height ( Figs 33B–C, E View FIGURE 33 ), but internal claws IV usually with primary spurs positioned higher (ca. 1/4–1- 3 of the claw branch) and strongly divergent from branches ( Fig. 33D View FIGURE 33 ). Sometimes, primary spurs IV developed similarly to those on claws I–III (compare Figs 33E and F View FIGURE 33 ).
Males. Not found.
Juveniles (i.e. the second instar; measurements and statistics in Table 9). Smaller than sexually mature individuals, but qualitatively identical. Gonopore absent.
Larvae (i.e. the first instar). Not found. Cuénot (1932) provided the body appendage formula A-Cd- Dd- E, where dorsal spines were significantly shorter than in adults.
Eggs. Up to five orange eggs per exuvia.
Phylogenetic position: Echiniscus spinulosus , with the current geographic coverage of sequenced populations, includes E. spiniger and forms an isolated clade with no close relatives (Figs 1–2). Importantly, the clade is remotely related to other numerous species with the spinulosus morphotype (trunk appendages in the form of spines, dorsal plate sculpturing consisting of dominant pores; see Discussion below).
Differential diagnosis: Echiniscus spinulosus is distinguished from other species of the spinulosus morphotype with dorsal appendages Cd and Dd significantly longer than lateral reduced spines/spicules on the basis of:
• E. tantulus Gąsiorek et al., 2020 , known only from Tanzania ( Bochnak et al. 2020): body size of sexually mature females (ca. 240–350 µm in E. spinulosus vs ca. 180–240 µm in E. tantulus ), body appendage configuration (A-(B)- C-Cd- D-Dd- E in E. spinulosus vs A-C-Cd- Dd- E in E. tantulus ), the presence of ornamentation (i.e. pores) in pedal plates (present in E. spinulosus vs absent in E. tantulus ), and claw heights (15.6–28.5 µm in E. spinulosus vs 9.8–15.0 µm in E. tantulus ).
Remarks: The allegedly cosmopolitan distribution of E. spinulosus ( McInnes 1994) is challenged with this redescription, as the previous records were again based on chaetotaxy, which is utterly misleading when used without details of dorsal plate sculpturing during identification of members of the spinulosus morphogroup. The verified geographic distribution embraces Western Palaearctic ( Fig. 40 View FIGURE 40 ), with definitely more abundant records from the Mediterranean, which could reflect xerophilous preferences of this species, often found on carbonate bedrock ( Dastych 1988). The examined historical material, freshly analysed populations, and, most importantly, the neotype population and population from Fontainebleau, exhibit a congruent morphology. Echiniscus carusoi Pilato, 1972 syn. nov. shares with E. spinulosus exactly the same type of dorsal plate sculpturing (including the two types of pores), ornamented pedal plates on all legs, and heteronych claws with identical spurs. Specimens with additional spicules (most individuals from the type series have the body appendage formula A-Cd- E) in the same positions as E. spinulosus corroborate the synonymising of the former, as it is only an aberrant population of E. spinulosus with reduced chaetotaxy. If the genetic uniformity of E. spiniger and E. spinulosus (Figs 1–2) is confirmed with a broader sampling, then E. spinulosus will be one of only two representatives of the spinulosus morphogroup (the other being E. scabrospinosus ) in continental Palaearctic, and characterised by a particularly large intraspecific variability.
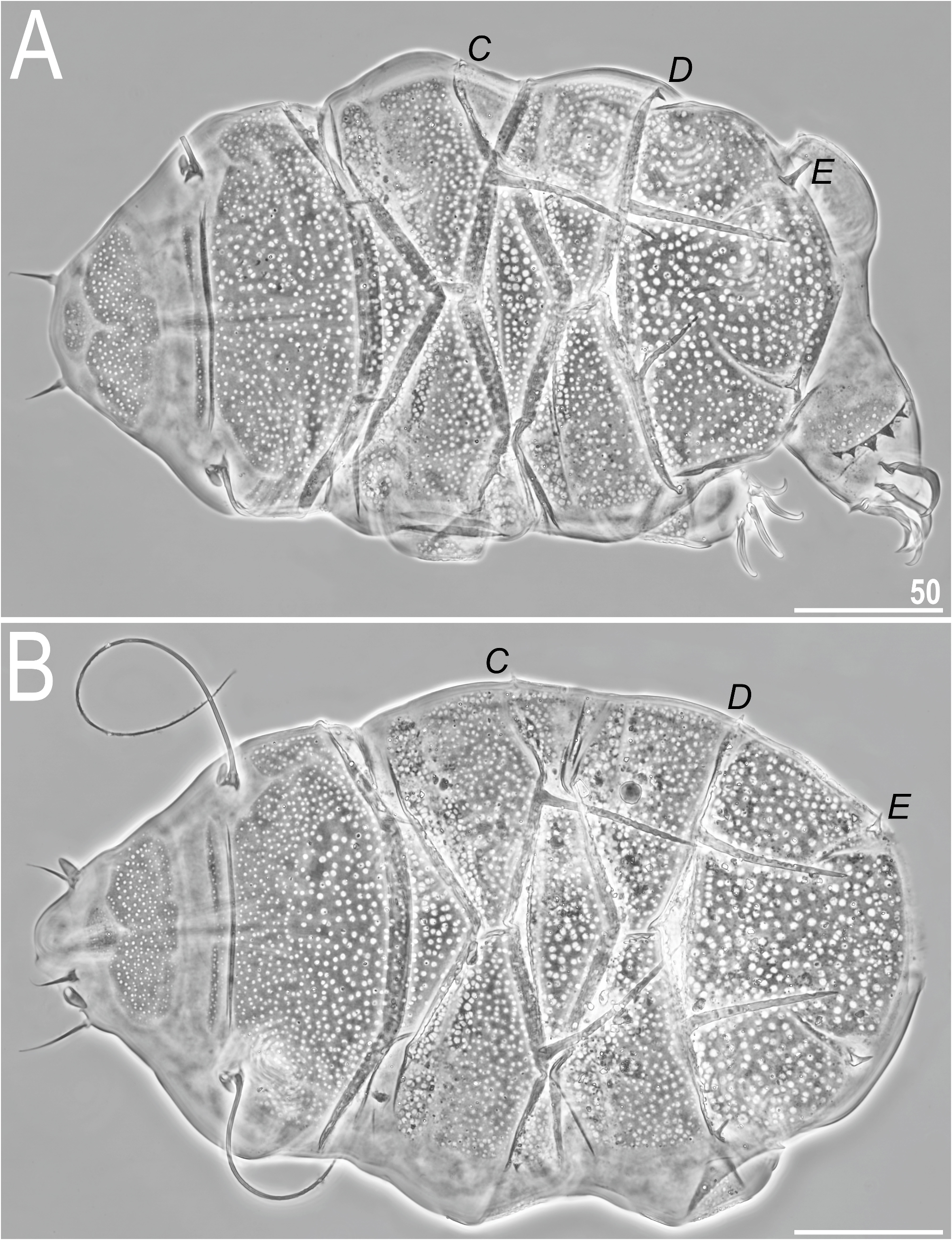



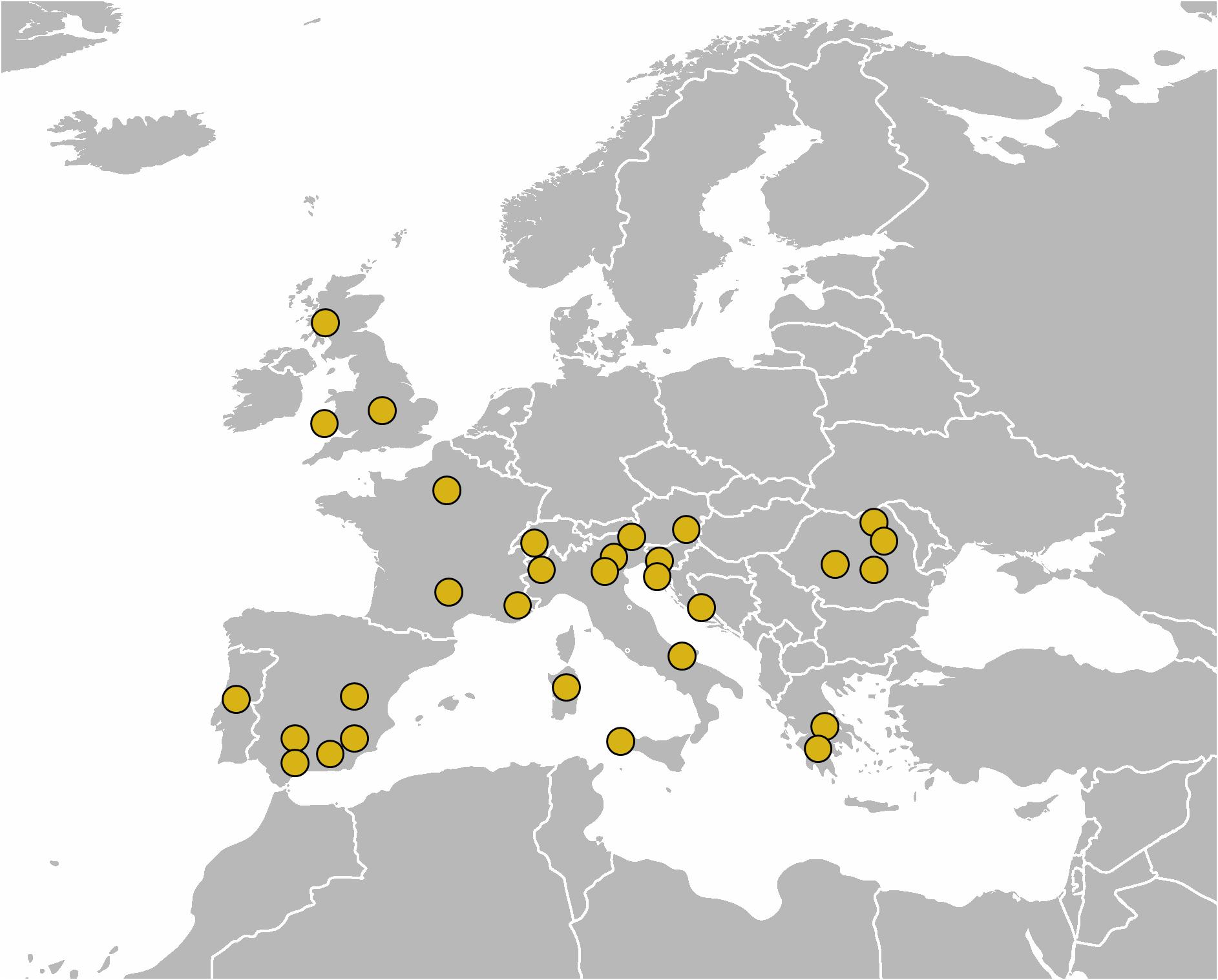
TABLE 8. Measurements [in µm] of selected morphological structures of adult females of Echiniscus spinulosus mounted in Hoyer’s medium. Abbreviations: sp—the proportion between the length of a given structure and the length of the scapular plate.
| CHARACTER | N | RANGE | MEAN | SD | Neotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | sp | µm | sp | µm | sp | µm | sp | ||||||
| Body length | 16 | 236 | – | 314 | 418 | – | 524 | 286 | 481 | 18 | 26 | 306 | 472 |
| Scapular plate length | 16 | 54.3 | – | 64.8 | – | 59.6 | – | 2.9 | – | 64.8 | – | ||
| Head appendage lengths | |||||||||||||
| Cirrus internus | 14 | 12.3 | – | 17.2 | 20.7 | – | 28.2 | 15.0 | 25.0 | 1.5 | 2.2 | 15.4 | 23.8 |
| Cephalic papilla | 16 | 8.6 | – | 10.5 | 14.5 | – | 17.8 | 9.4 | 15.8 | 0.5 | 0.8 | 9.7 | 15.0 |
| Cirrus externus | 15 | 20.2 | – | 29.1 | 37.0 | – | 49.8 | 25.5 | 42.7 | 2.6 | 3.8 | 29.1 | 44.9 |
| Clava | 11 | 6.7 | – | 9.1 | 10.3 | – | 15.2 | 7.7 | 13.0 | 0.6 | 1.3 | 6.7 | 10.3 |
| Cirrus A | 11 | 117.1 | – | 166.6 | 207.3 | – | 270.9 | 146.1 | 243.5 | 15.5 | 17.2 | 165.0 | 254.6 |
| Cirrus A /Body length ratio | 11 | 44% | – | 56% | – | 51% | – | 3% | – | 54% | – | ||
| Body appendage lengths | |||||||||||||
| Spine C | 10 | 2.2 | – | 5.2 | 3.6 | – | 9.4 | 3.4 | 5.7 | 0.9 | 1.7 | ? | ? |
| Spine Cd | 16 | 48.7 | – | 62.2 | 78.4 | – | 102.9 | 55.1 | 92.6 | 4.1 | 6.0 | 57.4 | 88.6 |
| Spine D | 15 | 2.5 | – | 10.0 | 4.3 | – | 17.7 | 6.6 | 11.0 | 1.8 | 3.2 | 7.1 | 11.0 |
| Spine Dd | 16 | 43.6 | – | 56.1 | 76.4 | – | 95.4 | 51.9 | 87.1 | 4.0 | 5.2 | 56.1 | 86.6 |
| Spine E | 16 | 5.4 | – | 13.4 | 9.5 | – | 20.7 | 8.7 | 14.6 | 2.1 | 3.1 | 13.4 | 20.7 |
| Spine on leg I length | 16 | 4.2 | – | 6.7 | 7.4 | – | 11.2 | 5.2 | 8.8 | 0.7 | 1.0 | 5.9 | 9.1 |
| Papilla on leg IV length | 16 | 4.5 | – | 5.5 | 7.6 | – | 9.1 | 4.9 | 8.3 | 0.3 | 0.4 | 5.5 | 8.5 |
| Number of teeth on the collar | 15 | 5 | – | 9 | – | 7.1 | – | 1.2 | – | 8 | – | ||
| Claw I heights | |||||||||||||
| Branch | 13 | 19.1 | – | 22.4 | 33.0 | – | 37.2 | 21.0 | 35.1 | 0.9 | 1.5 | 21.4 | 33.0 |
| Spur | 13 | 2.4 | – | 3.3 | 4.4 | – | 5.7 | 2.9 | 4.9 | 0.3 | 0.5 | 3.0 | 4.6 |
| Spur/branch height ratio | 13 | 12% | – | 16% | – | 14% | – | 1% | – | 14% | – | ||
| Claw II heights | |||||||||||||
| Branch | 15 | 16.2 | – | 21.5 | 28.4 | – | 37.2 | 19.8 | 33.2 | 1.4 | 2.2 | 20.8 | 32.1 |
| Spur | 15 | 2.3 | – | 3.4 | 4.0 | – | 5.6 | 2.9 | 4.9 | 0.3 | 0.5 | 3.4 | 5.2 |
| Spur/branch height ratio | 15 | 13% | – | 17% | – | 15% | – | 1% | – | 16% | – | ||
| Claw III heights | |||||||||||||
| Branch | 15 | 15.6 | – | 22.6 | 27.3 | – | 37.4 | 20.3 | 34.0 | 1.7 | 2.4 | 20.8 | 32.1 |
| Spur | 13 | 2.6 | – | 3.4 | 4.2 | – | 5.5 | 2.9 | 4.9 | 0.3 | 0.4 | 3.2 | 4.9 |
| Spur/branch height ratio | 13 | 12% | – | 17% | – | 14% | – | 1% | – | 15% | – | ||
| Claw IV heights | |||||||||||||
| Branch | 15 | 19.5 | – | 28.5 | 34.2 | – | 46.6 | 25.6 | 43.1 | 2.1 | 3.1 | 27.8 | 42.9 |
| Spur | 12 | 3.0 | – | 4.5 | 5.5 | – | 7.2 | 4.0 | 6.6 | 0.4 | 0.5 | 3.9 | 6.0 |
| Spur/branch height ratio | 12 | 12% | – | 17% | – | 15% | – | 1% | – | 14% | – | ||
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Echiniscus spinulosus ( Doyère, 1840 )
| Gąsiorek, Piotr & Vončina, Katarzyna 2023 |
Echiniscus carusoi
| Pilato 1972 |
Emydium spinulosum Doyère, 1840
| Doyere 1840 |