
Cyrtodactylus hoskini, Shea & Couper & Wilmer & Amey, 2011
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3146.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/71704301-FFF7-FFDA-FF36-F97FFCCDFB29 |
|
treatment provided by |
Felipe |
|
scientific name |
Cyrtodactylus hoskini |
| status |
sp. nov. |
Cyrtodactylus hoskini sp. nov.
( Figs. 19–20 View FIGURE 19 View FIGURE 20 )
Holotype. QM J86950 View Materials , female, Tozer's Gap , 205 m asl, Iron Range National Park (12º 43' 43" S 143º 11' 14" E) (P. Couper, A. Amey & L. Roberts, 15.ix.2008). GoogleMaps
Paratypes. QM J86926 View Materials –29, J86951 View Materials , Tozer's Gap , Iron Range National Park (12º 43' 43" S 143º 11' 14" E) GoogleMaps .
Diagnosis. A large Cyrtodactylus (SVL to 112 mm) with large, moderately projecting tubercles on the antebrachium, strongly developed dorsal tubercles in 19–24 longitudinal rows at the midpoint of the trunk (axilla-groin interval); 38–44 ventral scale rows at the same level, a continuous series of 41–48 enlarged femoroprecloacal scales extending from one knee to the other, each scale bearing a pore in males; mental with a posterior extension extending between postmentals; lips marbled or stippled with brown, dark dorsal bands on trunk usually three, with a narrow dark edge anteriorly and posteriorly, and a narrow vertebral extension both anteriorly and posteriorly; pale interspaces between dark body bands usually with a grey patch or bar laterally; basal tail bands a little wider than pale interspaces.
Description. Size large (subadult male 65 mm, n = 1; females 64–112 mm, mean = 100.0 mm, sd = 20.26, n = 5).
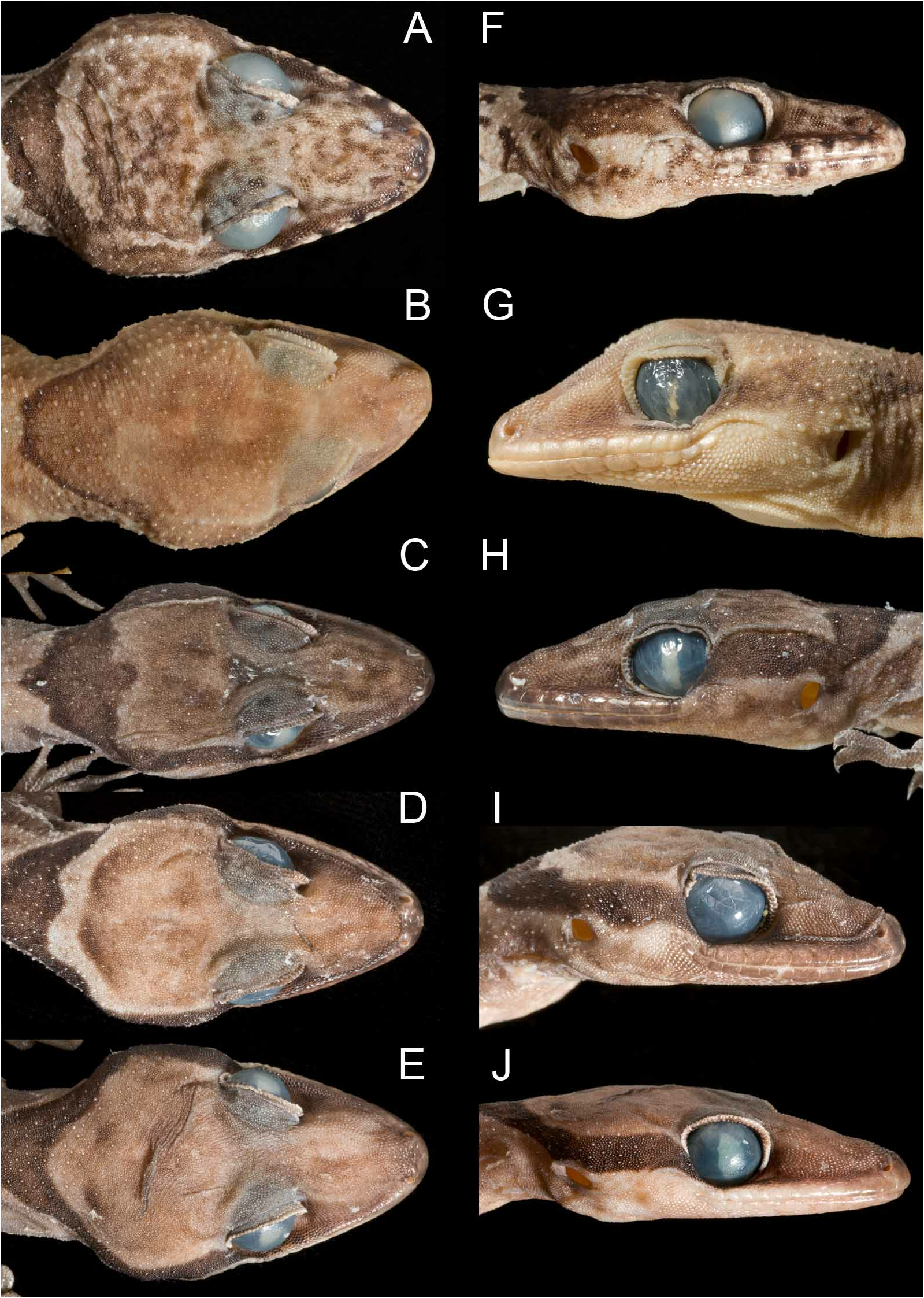
Head relatively long (HL/SVL 27.8–33.1%, mean = 30.0%; sd = 2.32, n = 6) and wide (HW/HL 62.7–69.8%, mean = 67.7%; sd = 2.87, n = 6), moderately depressed (HD/HL 34.0–37.1%, mean = 35.2%, sd = 1.16, n = 6), distinct from neck. Loreal region moderately inflated, canthus rostralis poorly defined. Interorbital region and top of snout concave, deepest and widest just anterior to level of rostral canthus of eye. Snout moderately long (SL/HL 36.3–39.5%, mean = 38.4%, sd = 1.18, n = 6; EN/SVL 27.4–29.4%, mean = 28.4%, sd = 0.76, n = 6), much longer than eye diameter (SL/EYE 142.6–176.1%, mean = 158.9%, sd = 12.74, n = 6), and a little longer than eye-ear interval (EE/HL 23.6–27.8%, mean = 26.0%, sd = 1.70, n = 6). Eye large (EYE/HL 22.4–25.9%, mean = 24.2%, sd = 1.33, n = 6), pupil vertical with crenated margin, forming about 3–4 low lobes along each edge of pupil. Supraciliaries in a double row, large, frill-like, well differentiated from adjacent more medial granules of the brow ridge, and largest anteriorly. Ear opening small (EAR/HL 7.7–10.2%, mean = 9.0%, sd = 0.84, n = 6), usually a little taller than long and slightly angled posterodorsally, but sometimes rounder. Rostral wider than high, height at centre less than that more laterally (except in QM J86951 View Materials ), dorsal part divided by a straight median groove that extends about ¼–⅓ the midline height of the scale, and fails to reach the oral margin. Two enlarged supranasals separated by usually a single, less enlarged internasal (two in QM J86929 View Materials ) that contacts the dorsal edge of the rostral shield. External nares circular, bordered by first supralabial, rostral, supranasal, nasal (extending into posterior part of nostril) and two smaller granular scales between nasal and first supralabial. Nares moderately separated (IN/HL 12.2–13.5%, mean = 12.9%, sd = 0.45, n = 6). Supralabials anteriorly large, distinct from adjacent loreal granules, 8–10 (mode = 8 (66.7%), mean = 8.7, sd = 1.03, n = 6) to level of mid-orbit, then inflecting dorsally and posteriorly, and becoming smaller, to gradually blend along rictal margin with adjacent small granules; supralabials separated from orbital margin by at least three rows of small granular scales at narrowest point. Mental wider than deep, with a strong median extension, equal to or slightly wider than rostral, and bordered posteriorly by a single elongate pair of large postmentals ( Fig. 10C View FIGURE 10 ). Infralabials anteriorly much larger than adjacent gular scales, becoming smaller posteriorly, 9–11 (mean = 10.0, sd = 0.89, n = 6). First infralabial with ventral border approximately ⅔ to fully contacting postmental. Subinfralabial scales anteriorly large, flattened, and polygonal, becoming smaller, more rounded and granular posteriorly and medially (towards gular area).
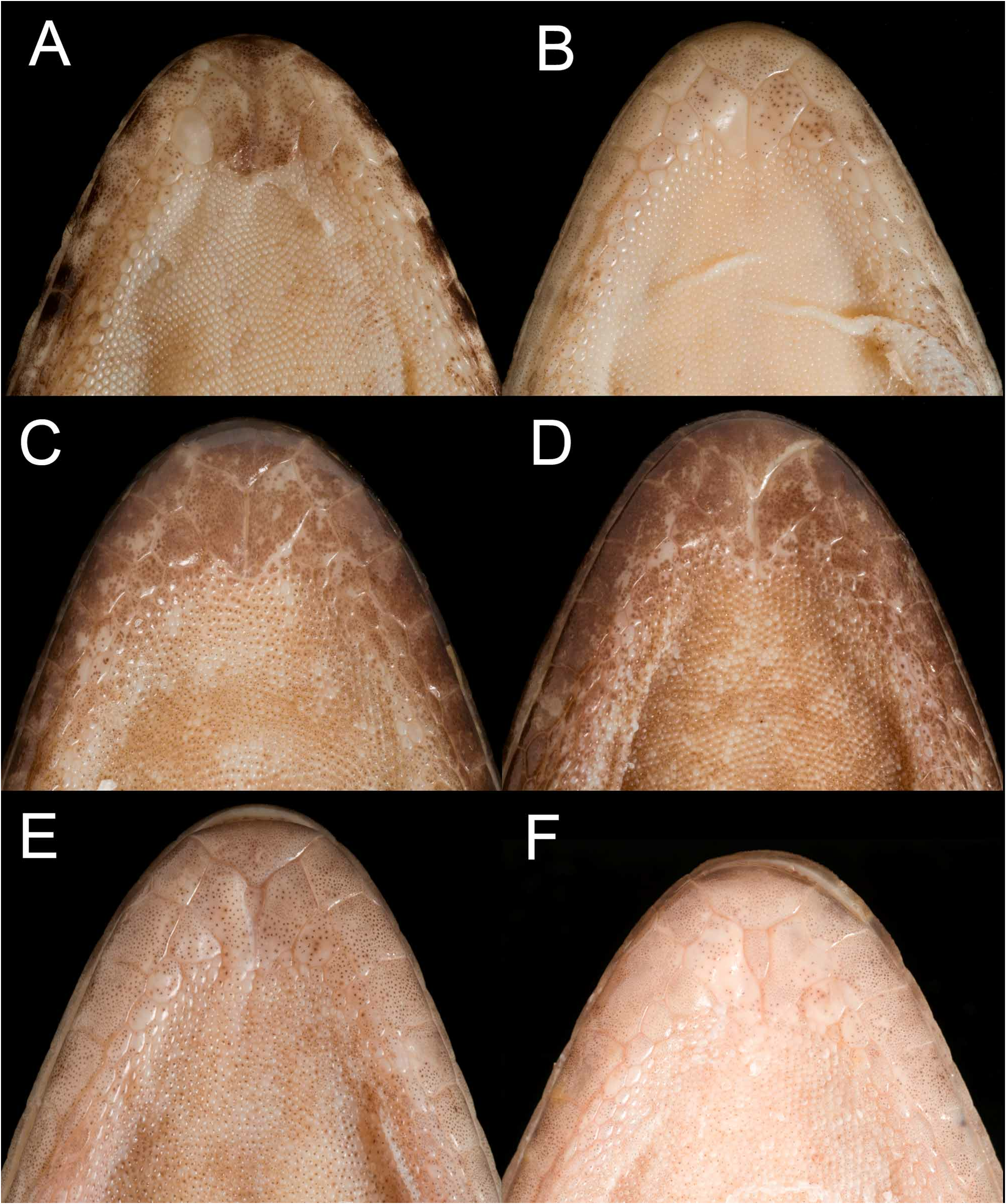
Body moderately robust (AGL/SVL 37.5–44.3%, mean = 41.0%, sd = 2.76, n = 6), with low, but distinct, ventrolateral skin folds approximately marking the transition between the enlarged flattened ventral scalation and the smaller, more rounded, granular lateral scalation. Scales on dorsum of head, body and limbs small, juxtaposed, rounded granules, with interspersed much larger tubercles. Granular scales finest over parietal region of head, becoming coarser over body, then larger, flatter and more polygonal on tail. On head dorsum ( Fig. 11C View FIGURE 11 ), tubercles commence on crown area and are small, widely separated and only slightly projecting, anteriorly, becoming larger, more numerous, more projecting and with a more conical, slightly posteriorly-tilted apex over nape. Tubercles on body dorsum larger again ( Fig. 12C View FIGURE 12 ), but with a more longitudinally ovoid base, often with a weak median keel, and relatively low on anterior body, but becoming more projecting posteriorly on body, over sacrum and onto tail base. Tubercles persist along tail, one to two whorls per segment, becoming lower and less differentiated until eventually losing their distinction by about the fourth or fifth dark band. Large tubercles on body dorsum separated by 2–3 smaller granular scales, those on head and nape more widely separated by several scales. Tubercles on body arranged in about 19–24 (mean = 21.7, sd = 1.63, n = 6) roughly longitudinal rows. Dorsum of brachium with juxtaposed small scales and relatively few larger tubercles; antebrachium with more imbricate, larger scales distally and over manus, and more numerous, large, low tubercles ( Fig. 13C View FIGURE 13 ). Dorsum of thigh and crus with small juxtaposed granules and numerous, low, densely packed, large tubercles ( Fig. 13H View FIGURE 13 ), only dorsum of pes with imbricate scales.
Laterally, tubercles commence over temporal region and in postinfralabial area, where they are noticeably larger than those of the head dorsum ( Fig. 11H View FIGURE 11 ), then along nape and body, where they are smaller and similarly protuberant to those dorsally, and along tail, commencing on tail base as prominent, conical scales, then rapidly losing differentiation by third dark tail band ( Fig. 14C View FIGURE 14 ).
Ventrally, gular scales small, rounded and juxtaposed, becoming larger, flatter and more imbricate over body venter, from clavicular region. Ventral scales at midbody, between ventrolateral skin folds 38–44 (mean = 39.8, sd = 2.23, n = 6). Ventral scales on brachium and antebrachium like gular scales. On ventral surface of thighs, but not on crus or in precloacal region, an abrupt junction between enlarged imbricate scales and much smaller scales posteriorly, enlarged scales 41–48 between distal extent on each thigh (mean = 44.5, sd = 2.74, n = 6). Ventral scales of tail base like those of body, most of tail venter with a single median series of very broad scales about four times the width of adjacent ventrolateral scales.
Precloacal and femoral pores present in males (n = 1), 48 in a single continuous row, arching shallowly anteriorly in precloacal region. No pubic groove. Usually two (three on left side of QM J86926 View Materials ) large, blunt-tipped, postcloacal spurs on ventrolateral surface of tail base.
Forelimbs and hindlimbs well-developed (FLL/SVL 15.5–16.5%, mean = 16.0%, sd = 0.34, n = 6; HLL/SVL 18.7–21.3%, mean = 19.6%, sd = 0.95, n = 6). Digits well-developed, reflected dorsally at proximal interphalangeal joint, and all bearing robust, strongly curved claws sheathed at the base by two scales. Subdigital lamellae expanded basally, beginning on pes over distal part of metatarsals and ending at point of reflection of toes, lamellae distal to this point not expanded. Lamellae under first toe 7–8 expanded (mean = 7.8, sd = 0.41) + 9–10 narrow (mean = 9.8, sd = 0.41), total 17–18 (mean = 17.7, sd = 0.52, n = 6). Lamellae under second toe 9–10 expanded (mean = 9.5, sd = 0.55) + 10–11 narrow (mean = 10.8, sd = 0.41), total 19–21 (mean = 20.3, sd = 0.82, n = 6). Lamellae under third toe 10–12 expanded (mean = 11.0, sd = 0.89) + 11–13 narrow (mean = 12.0, sd = 0.63), total 22–24 (mean = 23.0, sd = 0.89, n = 6). Lamellae under fourth toe 11–12 expanded (mean = 11.7, sd = 0.52) + 11–13 narrow (mean = 12.2, sd = 0.75), total 23–25 (mean = 23.8, sd = 0.98, n = 6). Lamellae under fifth toe 8–9 expanded (mean = 8.5, sd = 0.55) + 11–13 narrow (mean = 12.3, sd = 0.82), total 20–22 (mean = 20.8, sd = 0.75, n = 6). Relative lengths of digits on manus I<II<V<III<IV; on pes I<II<III=V<IV. Very slight traces of webbing between bases of fingers; weak webbing between bases of toes 2–3 and 3–4.
Tail a little longer than body (TL/SVL 133.0–147.7%, n = 2), narrow at base (TW/SVL 6.3–8.0%, mean = 7.3%, sd = 0.59, n = 6) and tapering evenly to a conical tip. Tail segments externally identifiable by straight scale junctions, segments about 7–8 scales long when counted to include tubercles. Cloacal sacs present in both sexes, external orifices just posterior to vent, laterally.
Colour in preservative. Dorsal ground colour pale grey-brown. Head dorsum ( Fig. 11C View FIGURE 11 ) usually coarsely mottled with mid to dark brown, bordered posterolaterally by a narrow pale zone which has a poorly defined anterior edge (except in juveniles, where the anterior edge is more clearly defined). Pale nape zone bordered posteriorly by a U-shaped dark chocolate coloured chevron on nape, widest vertebrally (with a short vertebral extension both anteriorly and posteriorly) and extending anteriorly over temporal region to eye, then moderately to weakly present over posterior lores and becoming increasingly diffuse as it extends to the nostril. Second broad dark transverse dorsal band over shoulders. Three dark bands over trunk, narrowing on lateroventral surfaces and dissipating over lower flanks. Usually a dark band over hips (not present in QM J86950 View Materials and QM J86928 View Materials ; both specimens with a narrow dark blotch at the limb insertions, the latter specimen also has a small dark blotch on the vertebral line). Tail with dark bands over most of length, but these become increasingly obscure on the distal portion. When they can be counted to the distal end of tail, dark tail bands 13–15 (n = 2). On nape and body, dark bands usually slightly wider than pale interspaces, and with abrupt edges with a short vertebral extension both anteriorly and posteriorly; pigmentation darkest along band margins, leaving centre of bands a little lighter. Bands on tail ( Fig. 14C View FIGURE 14 ) of similar width to body bands, and anteriorly 1½ to twice the width of the pale interspaces. Pale interspaces of body usually with a few dark smudges and some indication of a dark bar that extends dorsoventrally down the flanks.
Upper and lower lips ( Fig. 11H View FIGURE 11 ) generally greyish-brown. Dorsum of forelimbs and hindlimbs mid-brown and mottled; in some specimens obscure darker bars are present on the thighs.
Entire ventral surface generally pale with some greyish brown mottling which is most prominent on the mental, the gular region and the jaw articulation.
Description of holotype. The holotype of C. hoskini is a mature-sized female, with the following character states of those variable for the taxon: SVL 107.5 mm, AGL 45 mm, TL 143 mm, TW 8.6 mm, HL 29.9 mm, HW 20.8 mm, HD 11.1 mm, IN 3.8 mm, SL 11.8 mm, EN 8.5 mm, EYE 6.7 mm, EE 8.3 mm, EAR 2.7 mm, FLL 16.9 mm, HLL 21.2 mm, lamellae below digits I–V 8+10, 10+11, 12+12, 12+12, 9+13 respectively, supralabials 8, infralabials 9, rows of dorsal tubercles 22, transventral rows 40, femoroprecloacal scales 41, dark hip band present, and dark tail bands 15.
Etymology. Named for Dr Conrad Hoskin (b. 1976) whose work on rainforest frogs and reptiles played an important role in recognizing the significance of rock landscapes (lithorefugia) in preserving ancient rainforest lineages (see Couper & Hoskin 2008); a concept that is well illustrated by the distribution of Australian Cyrtodactylus spp.
Distribution. The western edge of Iron Range, north Queensland ( Fig. 21 View FIGURE 21 ). In addition to the type locality, the species has been photographed in the Garraway Creek area (12º 42' 50" S 143º 10' 26" E), a locality approximately 2 km north of the type locality (C. Hoskin, pers. comm.), and from the William Thompson Range (at the point where Fall Creek drops out of the William Thompson Range, just west of the Pascoe River Crossing, 12º 55' 27" S 143º 01' 30" E; K. Aland, pers. comm.), about 29 km to the south-west ( Fig. 20B View FIGURE 20 ).
Conservation status. This species has a very small known distribution that is unlikely to be much more extensive. We have no clear understanding of population size but expect it to be small, given the very small area of occupancy and the low genetic diversity of this species on current sampling. The large adult size and colourful pattern of this species, together with its ready collection from accessible rock outcrops, could lead to it being targeted by illegal collection from the wild for the pet trade. These factors fulfil the IUCN criteria for a Vulnerable listing (criteria D1, VU D2).
Comparison with other species ( Table 6). Cyrtodactylus hoskini is genetically most similar to C. tuberculatus and C. mcdonaldi , differing from them by average sequence divergences of 12.12% and 11.79% respectively ( Table 2). In comparison to C. tuberculatus , C. hoskini has more numerous transverse ventral scales (38–44 vs 24–37) and precloacal scales (41–48, mean = 44.5 vs 34–46, mean = 39.8), a narrower, flatter head (HW/HL 62.7–69.8% vs 66.5–88.0%; HD/HL 34.0–37.1% vs 35.3-57.5%), with a less well-developed postocular region (EN/HL 23.6–27.8% vs 25.8–36.9%), and is generally less tuberculate than C. tuberculatus , with the postlabial area barely tuberculate (vs strongly tuberculate), the crus with only small, low tubercles (vs strongly tuberculate), and caudal tubercles absent by or at the second pale band (vs continuing to the third or fourth pale band or more, distally). In coloration, C. hoskini has a generally less strongly mottled head dorsum (vs usually strongly mottled), strong anterior extensions of dark body bands along the vertebral line (barely present in a few individuals of C. tuberculatus , but usually absent), and a dark bar in the pale interspaces ventrolaterally (absent in C. tuberculatus , or represented by scattered rounded spots).
Cyrtodactylus hoskini has more numerous transverse ventral scales than C. mcdonaldi (38–44 vs 28–41), more numerous enlarged femoroprecloacal scales (41–48 vs 37–45), a longer tail (TL/SVL 133.0–147.7% vs 117.0–128.7%), a narrower and flatter head (HW/HL 62.7–69.8% vs 65.0–78.9%; HD/HL 34.0–37.1% vs 35.8–43.4%), with a less well-developed temporal region (EE/HL 23.6-27.8% vs 27.6-33.1%), and slightly longer hindlimbs (HLL/SVL 18.7–21.3% vs 16.5–19.2%). The subdigital lamellae counts are at the upper end of the range for C. mcdonaldi . The femoroprecloacal pores are in a single unbroken row (vs broken into three segments by unpored scales). The dark bands on body and nape have a strong anterior extension along the vertebral line (vs absent), the pale interspaces have a dark bar ventrolaterally (vs usually no dark markings), and the dark dorsal bands have strong anterior and posterior edges (usually only a pronounced dark posterior edge in C. mcdonaldi ).
Cyrtodactylus hoskini differs from C. klugei and C. robustus in having many fewer femoroprecloacal pores (48 vs 66 or more) in males. The difference in number of pores is mirrored by the number of scales bearing them, which are countable in both males and females (41–48 ( C. hoskini ) vs 66 or more). In comparison to C. klugei , C. hoskini has three dark bands across the trunk (vs usually two), and has vertebral extensions of the dark body bands (vs absent). It is less strongly tuberculate than C. robustus , is smaller (SVL to 105 mm vs 161 mm) and has a narrower head (HW/HL 62.7–69.8% vs 69–84%; Kraus 2008), and further differs from it in the presence of a dark band across the hips (vs usually lacking). It also lacks the bright orange cloaca of C. robustus .
For comparisons with C. adorus and C. pronarus , see the descriptions of those species.
Comments. The discovery of a Cyrtodactylus species at Iron Range, the stated type locality for Cyrtodactylus abrae Wells, 2002 , requires revisiting the status of that nominal taxon. Couper et al. (2004) recommended treating the name as unavailable, due to the lack of a type specimen in the original description. However, if a neotype were to be designated, the name could be validated. The limited description of C. abrae , which appears to be based on previous accounts of a "rainforest" form of Cape York Cyrtodactylus presented by Wilson and Knowles (1988), in turn seemingly based on the published photograph by Cogger (1975), clearly does not represent the taxon we have described from Iron Range. Wells (2002) defined C. abrae as being differentiated from C. tuberculatus by its possession of fewer dark body bands (four, corresponding to two in our definition), and tail bands (seven). Cyrtodactylus hoskini , the only Cyrtodactylus that we have evidence of at Iron Range, has the same number of dark body bands (three) as C. tuberculatus , and none of the Australian Cyrtodactylus species have as few as seven dark tail bands or two dark body bands between axilla and groin. Wells (2002) also noted that C. abrae can attain a SVL of 160 mm, considerably larger than any other Australian Cyrtodactylus . Hence, the possibility exists that the description of C. abrae is based on a non-Australian taxon with incorrect type locality, and on current knowledge, it is not possible to nominate a neotype that is both from a locality close to the nominal type locality as well as according with the nominal diagnosis of the taxon. Because of the considerable risk to destabilization of nomenclature by having this name potentially become a senior synonym of any one of the numerous Cyrtodactylus species described since 2002 through nomination of a neotype, and the lack of sufficient detail in the description (or in the photograph provided by Cogger (1975) that seems to have been the basis for that description) to clearly assign it to any one of the numerous banded Cyrtodactylus species , we choose to stabilize nomenclature by nominating as neotype of Cyrtodactylus abrae the larger syntype of Cyrtodactylus pulchellus , Natural History Museum, London (BMNH) xxii.91a, from Singapore. This action accords with the sparse detail that is available in the morphological and coloration description of C. abrae by Wells (2002), and relegates the name to synonymy. As we have demonstrated, it is not possible to nominate a neotype that is as close as possible to the nominal type locality of C. abrae , consistent with Article 75.3.6 of the Code of Zoological Nomenclature ( ICZN 1999), because none of the Australian Cyrtodactylus are morphologically consistent with the description of C. abrae . To comply with Article 75.3.3 and 75.3.5, relevant to validation of neotype designations, we illustrate this specimen (the larger syntype of C. pulchellus ) in dorsal view ( Fig. 22 View FIGURE 22 ).
Natural history. The type series of C. hoskini was collected amongst large granite boulders in predominantly open forest interspersed with patches of rainforest ( Fig. 23A View FIGURE 23 ). The nearby Garraway Creek site is immediately adjacent to the type locality for the microhylid frog Cophixalus kulakula , which is described and illustrated by Hoskin and Aland (2011) as a boulder field, with festooning ferns vines and umbrella trees. The William Thompson Range population inhabits granite boulders in outcrops and along creek and gully lines in savannah woodland that shows evidence of regular burns (K. Aland, pers. comm.).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |