
Corydoras areio Knaack, 2000
|
publication ID |
https://doi.org/ 10.1590/1982-0224-2020-0088 |
|
persistent identifier |
https://treatment.plazi.org/id/03B787AD-AA41-FFC1-1566-106389D64B8A |
|
treatment provided by |
Felipe |
|
scientific name |
Corydoras areio Knaack, 2000 |
| status |
|
Corydoras areio Knaack, 2000 View in CoL
( Figs. 1 View FIGURE 1 , 2A View FIGURE 2 , 3A View FIGURE 3 , 4 View FIGURE 4 )
Corydoras areio Knaack, 2000:45 , 47, 49, 51-53 (original description; type-locality: “ Brasilien, Mato Grosso (östlich von Cuiaba), in Fliessgewässern des Córr. Areio- Systems” [= Brazil, Mato Grosso (east of Cuiabá), in the flowing waters of the córrego Areio basin]). —Reis, 2003:295 (listed). —Fuller, Evers, 2005:70 (photo in life; presence in the aquarium hobby; córrego Areio basin). —Britski et al., 2007:154–155 (identification key; brief description). —Ferraris, 2007:113 (listed).
Corydoras aurofrenatus (non Eigenmann, Kennedy, 1903). —Britto, 2003: 153 (partim; specimens from MZUSP 36720).
Diagnosis. Corydoras areio can be distinguished from its congeners, except for the species within lineage 1 sensu Alexandrou et al. (2011), by the presence of the following features: temporal sensory canal in sphenotic with two pores anteriorly to the branch that gives rise to infraorbital canal (vs. with a single pore), upper tooth plate of branchial arch with three or four series of teeth (vs. two series of teeth), area at the corner of the mouth, ventral to the maxillary barbel, with a fleshy flap (vs. fleshy flap absent). Corydoras areio can be distinguished from the species belonging to the lineage 1, except for C. acutus Cope, 1872 , C. cervinus Rössel, 1962 , C. maculifer Nijssen, Isbrücker, 1971 , C. oxyrhynchus Nijssen, Isbrücker, 1967 , C. saramaccensis Nijssen, 1970 , C. vittatus Nijssen, 1971 and C. zawadzkii Tencatt & Ohara, 2016 , by the absence of large patches of black pigmentation on the body (vs. presence in the remaining congeners). Corydoras areio is diagnosed from C. acutus , C. cervinus , C. maculifer , C. vittatus and C. zawadzkii by the presence of blotches on flanks not aligned in longitudinal series (vs. aligned in longitudinal series, sometimes forming continuous bands). Corydoras areio can still be distinguished from C. vittatus plus C. saramaccensis by the absence of a dark brown or black stripe transversally crossing the eye (vs. presence of such stripe, forming the typical mask-like blotch). Corydoras areio also differs from C. cervinus by having infraorbital 2 with relatively wider posterior laminar expansion (vs. infraorbital 2 with relatively narrower posterior laminar expansion). Finally, C. areio is diagnosed from C. oxyrhynchus by the absence of conspicuous concentration of dark brown or black chromatophores on anterior portion of dorsal fin, especially close to dorsal-fin spine insertion (vs. presence of such pattern).
Description. Morphometric data are presented in Tab. 1. Head compressed with convex dorsal profile, roughly triangular in dorsal view. Snout conical conspicuously pointed. Head profile slightly concave from tip of snout to anterior nares, ascending straight to slightly convex from this point to anterior portion of parieto-supraoccipital; slightly convex from this point to tip of posterior process of parieto-supraoccipital. Profile slightly convex along dorsal-fin base. Postdorsal-fin body profile concave to adipose-fin spine, concave from this point to caudal-fin base. Ventral profile of body nearly straight from isthmus to pectoral girdle, and slightly convex from this point until pelvic girdle. Profile nearly straight from pelvic girdle to base of first anal-fin ray, roughly concave until caudal-fin base. Body roughly elliptical in cross section at pectoral girdle, gradually becoming more compressed toward caudal fin.
Eye rounded, located dorsolaterally on head. Orbit delimited anteriorly by lateral ethmoid, anterodorsally by frontal, posterodorsally by sphenotic, posteriorly by infraorbital 2, and ventrally by infraorbital 1. Anterior and posterior nares close to each other, only separated by flap of skin. Anterior naris tubular. Posterior naris close to anterodorsal margin of orbit, separated from it by distance similar to naris diameter. Mouth small, subterminal, width nearly equal to bony orbit diameter. Maxillary barbel long in size, reaching anteroventral limit of gill opening. Outer mental barbel slightly longer than maxillary barbel. Area at corner of mouth, ventral to maxillary barbel, with reduced fleshy flap. Inner mental barbel fleshy, base of each counterpart slightly separated from each other. Small rounded papillae covering entire surface of all barbels, upper and lower lips, snout and isthmus.
Mesethmoid long with anterior tip well developed, larger than 50% of bone length (see Britto, 2003:123, character 1, state 0; fig. 1A), and posterior portion relatively narrow, entirely covered by thin layer of skin. Middle portion of mesethmoid with well-developed lateroventral process; region of process with width similar to width of posterior portion of mesethmoid. Nasal capsule delimited anterodorsally by mesethmoid, anteriorly and ventrally by lateral ethmoid, and posteriorly and dorsally by frontal. Nasal slender, laterally curved, inner margin laminar, and mesial border contacting frontal and mesethmoid. Lateral ethmoid conspicuously expanded anteriorly, with anterodorsal expansion contacting only mesethmoid, and anteroventral expansion connected to lateroventral process of mesethmoid. Frontal elongated, strongly narrow, width clearly smaller than half of its entire length; anterior projection short, size smaller than nasal length. Frontal fontanel large, conspicuously slender, posterior tip extension markedly entering anterior margin of parieto-supraoccipital. Sphenotic somewhat trapezoid, contacting parieto-supraoccipital dorsally, compound pterotic posteriorly, second infraorbital ventrally and frontal anteriorly ( Fig. 2A View FIGURE 2 ). Compound pterotic roughly pipe-shaped, with posteriormost portion contacting first lateral-line ossicle, posteroventral margin contacting cleithrum, and anteroventral margin contacting opercle and infraorbital 2, and posterior expansion almost entirely covering lateral opening of swimbladder capsule, leaving slender area on its dorsal margin covered only by thick layer of skin ( Fig. 2A View FIGURE 2 ). Parieto-supraoccipital wide, posterior process long and contacting nuchal plate and region of contact between posterior process and nuchal plate covered by thick layer of skin.
Two laminar infraorbitals with minute odontodes. Infraorbital 1 large, ventral laminar expansion ranging from moderately- to well developed; anterior portion with well-developed laminar expansion, reaching to or slightly surpassing anterior margin of nasal capsule; inner laminar expansion strongly reduced ( Fig. 2A View FIGURE 2 ). Infraorbital 2 small, widened, with posterior laminar expansion well developed, and posteroventral margin contacting posterodorsal ridge of hyomandibula, posterodorsal edge contacting sphenotic and compound pterotic; inner laminar expansion poorly developed ( Fig. 2A View FIGURE 2 ). Posterodorsal ridge of hyomandibula close to its articulation with opercle conspicuously slender, exposed, reduced and bearing small odontodes. Dorsal ridge of hyomandibula between compound pterotic and opercle covered by thick layer of skin. Interopercle covered by thin layer of skin, subtriangular, anterior projection well-developed. Preopercle relatively slender, elongated, minute odontodes sparse on external surface. Opercle dorsoventrally elongated, width equal to or smaller than half of entire length; free margin slightly convex, without serrations and covered by small odontodes.
Four branchiostegal rays decreasing in size posteriorly. Hypobranchial 2 somewhat triangular, tip ossified and directed towards anterior portion, posterior margin cartilaginous, and ossified portion conspicuously well developed, its size three times or more than cartilaginous portion. Five ceratobranchials with expansions increasing posteriorly; ceratobranchial 1 generally with strongly reduced process on anterior margin of mesial portion; ceratobranchial 3 with continuous laminar expansion on postero-lateral margin; ceratobranchial 5 toothed on posterodorsal surface, with 22 to 26 (2) teeth aligned in one row. Four epibranchials with similar size; epibranchial 2 slightly larger than others, with small pointed process on laminar expansion of posterior margin; epibranchial 3 with mesially-curved uncinate process on laminar expansion of posterior margin. Two wide pharyngobranchials (3 and 4), pharyngobranchial 3 with triangular laminar expansion variably notched on posterior margin. Upper tooth plate oval, 42 to 51(2) teeth roughly aligned in three or four rows on posteroventral surface.
Lateral-line canal reaching cephalic laterosensory system through compound pterotic, branching twice before reaching sphenotic: pterotic branch, with single pore, preoperculomandibular branch conspicuously reduced, with single pore opening close to postotic main canal, postotic main canal becoming widened just posterior to pterotic branch. Sensory canal continuing through compound pterotic, reaching sphenotic as temporal canal, which splits into two branches: one branch giving rise to infraorbital canal, other branch connecting to frontal through supraorbital canal, with one and two pores, respectively. Supraorbital canal branched, running through nasal bone. Epiphyseal branch relatively long, pore opening close to frontal fontanel. Nasal canal with two or three openings, first on posterior edge, second, when present, on posterolateral portion and generally fused with first pore, and third on anterior edge. Infraorbital canal running through entire infraorbital 2, extending to infraorbital 1 and opening into two pores. Preoperculomandibular branch giving rise to preoperculomandibular canal, which runs through entire preopercle with three openings, leading to pores 3, 4, and 5, respectively.
Dorsal fin subtriangular, located just posterior to third dorsolateral body plate. Dorsal-fin rays II,7 (1), II,8 (39), II,9 (1), posterior margin of dorsal-fin spine with seven to 10 poorly-developed serrations directed towards tip of spine, serrations arranged on distal half of posterior margin; small odontodes on anterior and lateral surfaces of spine. Nuchal plate well developed, almost entirely exposed, with minute odontodes. Spinelet short, spine moderately developed, adpressed distal tip slightly surpassing posterior origin of dorsal-fin base, and anterior margin with small odontodes. Pectoral fin roughly triangular, its origin just posterior to gill opening. Pectoral-fin rays I,9 (15), I,10 (26), posterior margin of pectoral spine with 16 to 17 with moderately- to well-developed conical serrations along its entire length, most serrations directed towards pectoral-fin origin, and some serrations perpendicularly directed or directed towards tip of spine; small odontodes on anterior, dorsal and ventral surfaces of spine ( Fig. 3A View FIGURE 3 ). Anteroventral portion of cleithrum exposed; posterolateral portion of scapulocoracoid exposed; small odontodes on exposed areas. Pelvic fin oblong, located just below third ventrolateral body plate, and at vertical through first branched dorsal-fin ray. Pelvic-fin rays i,5. Adipose fin roughly triangular, separated from base of last dorsal-fin ray by generally six dorsolateral body plates. Anal fin subtriangular, located just posterior to 12 th ventrolateral body plates, and at vertical through anterior margin of adipose-fin spine. Anal-fin rays ii,5 (3), i,7 (1), ii,6 (16). Caudal fin bilobed, markedly furcated, with dorsal lobe slightly larger than ventral lobe. Caudal-fin rays i,12,i, generally four dorsal and ventral procurrent rays.
Two to four laterosensory canals on trunk. First ossicle tubular, second ossicle laminar and the remaining encased in third, fourth and fifth dorsolateral body plate, respectively. Body plates with minute odontodes scattered over exposed area, conspicuous line of odontodes confined on posterior margins. Dorsolateral body plates 23 (30), 24 (9). Ventrolateral body plates 20 (8), 21 (31). Dorsolateral body plates along dorsal-fin base 6. Dorsolateral body plates between adipose- and caudal-fin 7 (13), 8 (7). Preadipose platelets 1 (1), 2 (6), 3 (26), 4 (5), 5 (3). Small platelets covering base of caudal-fin rays. Small platelets disposed dorsally and ventrally between junctions of lateral plates on
posterior portion of caudal peduncle. Anterior margin of orbit, above the junction of frontal and lateral ethmoid, ventral margin of nasal capsule and dorsal surface of snout with small, irregular platelets bearing odontodes. Ventral surface of head and trunk densely covered by small irregular platelets bearing odontodes.
Vertebral count 22 (2). Ribs 5 (2), first pair conspicuously large, its middle portion closely connected to first ventrolateral body plate. Complex vertebra moderately developed.
Corydoras areio Corydoras aurofrenatus
Low-High Mean±SD Low-High Mean±SD
Standard length (mm) 33.4–42.1 37.6±2.2 29.7–47.6 40.4±5.2
Percentages of standard length
Depth of body 34.0–37.4 35.2±1.0 32.1–37.8
35.0 ± 1.5
Predorsal distance 46.9–51.9 49.4±1.2 45.9–50.8
48.3 ± 1.1
Prepelvic distance 46.9–50.0 48.2±0.9 45.4–50.3
47.6 ± 1.2
Preanal distance 78.2–82.4 80.8±1.2 76.9–82.5
79.7 ± 1.6
Preadipose distance 80.7–85.8 83.3±1.3 80.2–84.8
82.1 ± 1.2
Length of dorsal spine 19.6–23.2 21.2±1.0 18.2–23.9
20.9 ± 1.4
Length of pectoral spine 20.5–26.7 23.3±1.8 19.1–24.0
21.8 ± 1.3
Length of adipose-fin spine 8.1–11.6 9.2-0.8 7.4–11.4
9.4 ± 1.0
Depth of caudal peduncle 13.1–14.6 14.0±0.4 12.8–15.4
14.0 ± 0.7
Length of dorsal-fin base 16.2–18.7 17.6±0.7 15.4–20.6
18.1 ± 1.1
Dorsal to adipose distance 17.7–21.7 19.6±0.9 15.9–21.6
18.6 ± 1.5
Maximum cleithral width 24.5–26.2 25.4±0.5 23.3–27.0
25.3 ± 1.1
Head length 40.3–44.5 42.2±1.1 38.4–43.4
41.4 ± 1.3
Length of maxillary barbel 18.9–23.6 21.0±1.4 13.2–22.4
18.9 ± 2.4
Percentages of head length
Head depth 72.7–81.9 77.4±1.9 69.8–82.8
77.7 ± 2.8
Least interorbital distance 21.8–25.2 23.4±1.0 21.7–26.9
24.3 ± 1.2
Horizontal orbit diameter 20.9–23.2 22.0±0.7 18.3–22.5
20.2 ± 1.3
Snout length 39.1–48.7 45.0±2.4 38.6–45.6
42.7 ± 1.9
Least internarial distance 9.3–14.7 12.6±1.4 8.3–13.0 10.9 ± 1.5
Coloration in alcohol. Overall color of body in Fig. 1 View FIGURE 1 . Ground color of body yellow. Top of head dark brown. Region just above posterodorsal margin of orbit with nearly straight, horizontally elongated dark brown or black blotch, forming eyebrow-like marking, blotch slightly arched, following outline of orbit in some specimens, variably diffuse or absent. Dorsal surface of snout with conspicuous concentrations of dark brown or black chromatophores, variably forming rounded or irregular blotches generally diffuse. Ventral region of infraorbital 1 with conspicuous concentration of dark brown or black chromatophores, with pigmentation extending ventrally in anterior- and posterior-most infraorbital 1 edges in some specimens. Opercle with border and middle portion yellow, remaining area with conspicuous concentration of dark brown or black chromatophores. Cleithrum with conspicuous concentrations of dark brown or black chromatophores on its dorsolateral surface, variably forming irregular small blotches; blotches absent or diffuse in some specimens. Dorsal series of diffuse dark brown or black blotches, first on anterior portion of dorsal-fin base, second on posterior portion of dorsal-fin base, third on adipose-fin base and the last one on posterior portion of caudal peduncle. First blotch conspicuously circular, remaining blotches irregular or rounded. Dorsolateral body plates with relatively small rounded, irregular or elongated dark brown blotches. Ventrolateral body plates generally unspotted, dark brown rounded, irregular or elongated blotches close to midline of flank in some specimens. Last dorso- and ventrolateral body plates with conspicuous concentration of dark brown or black chromatophores, forming generally diffuse, irregular, transversally elongated blotch; some specimens with conspicuous blotch. Spots on flanks faded in few specimens. Dorsal-fin with conspicuous concentration of dark brown or black chromatophores, generally more evident on rays, forming small spots; some specimens with diffuse spots. Pectoral and pelvic fins with dark brown or black chromatophores, generally more concentrated on rays and not forming spots. Adipose fin with dark brown or black chromatophores, generally more concentrated on spine, especially on its distal two-thirds and ventral portion of its origin. Anal fin with conspicuous concentration of dark brown or black chromatophores, generally more evident on rays of its middle portion, forming small spots; spots roughly aligned transversally and generally diffuse. Middle portion of caudal-fin base with small and diffuse dark brown or black dot. Caudal fin with conspicuous concentrations of dark brown or black chromatophores, generally more evident on rays, forming blotches roughly aligned transversally in four to ten slender bars; diffuse bars in some specimens.
Coloration in life. Similar to color pattern of preserved specimens but with lighter ground color of body, and with greenish yellow iridescent coloration ( Fig. 4 View FIGURE 4 ). Eyebrow-like blotch generally more evident. Spots faded in some specimens.
Sexual dimorphism. The presence of lanceolate genital papilla is a common feature in males of all Corydoradinae species (see Nijssen, Isbrücker, 1980b; Britto, 2003). Additionally, the males present a fibrous tissue on pectoral spine, which is generally covered by hypertrophied odontodes, and also a segmented filament on the tip of the spine, as illustrated for C. solox Nijssen & Isbrücker, 1983 (see Nijssen, Isbrücker, 1983:81, fig. 10g).
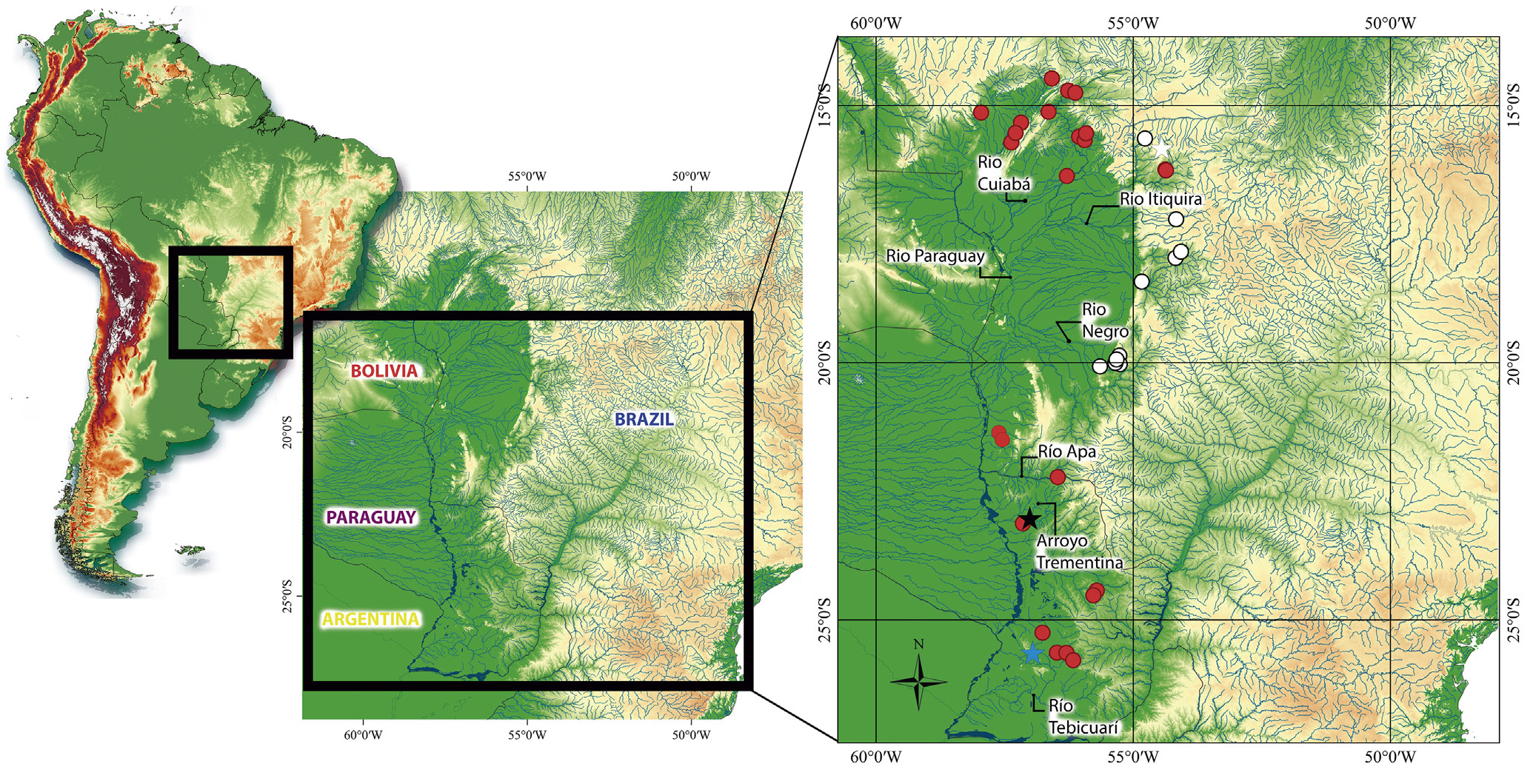
Geographical distribution. Corydoras areio is known from the rio Piquiri (rio
Itiquira) and the ribeirão Parnaíba, tributaries of the rio São Lourenço basin, State of
Mato Grosso, and also from the rio Negro (rio Taboco) and rio Taquari basins, State of
Mato Grosso do Sul, Brazil ( Fig. 5 View FIGURE 5 ).
Ecological notes. In the rio Taboco and rio Taquari basins, Corydoras areio was
generally found inhabiting small streams ( Fig. 6 View FIGURE 6 ), mainly associated with deep, lentic
habitats during the day, ranging from about 50 cm to 1 m depth, with substrate
predominantly composed of sand. During day, the species displays a more active
behavior, swimming away at the sign of any movement, even from outside of the water.
On the other hand, the species can be easily observed and captured in shallow beaches
of streams (about 10 cm depth) during the night, where they stay nearly motionless. At
the beaches of the ribeirão dos Veados ( Fig. 6A View FIGURE 6 ), the species was observed in syntopy
with C. polystictus and Corydoras aff. polystictus . The species was rarely captured in the
main channel of the rio Taboco ( Fig. 6B View FIGURE 6 ), where it is also associated with the sandy
substrate of the river banks. In the rio Taboco basin, C. areio occurred in syntopy with
Corydoras aff. aeneus (Gill, 1858) . In most collecting sites, C. areio specimens were
observed burying themselves in the sandy substrate, especially during capture attempts.
Another interesting feature of C. areio observed in natural habitat is the presence of
translucid trunk in smaller specimens (up to 30.0 mm SL), which possibly enhances their camouflage on sand. Additionally, it was possible to observe signs of severe deforestation in the region of both river basins, basically for agricultural and cattle raising purposes, which reflects the high levels of siltation in most of the local water bodies ( LFCT pers. obs.).
Remarks. An interesting point concerning Corydoras areio is its exact type-locality,
since Knaack (2000) did not provide an exact locality, pointing that the species was
captured in streams of the “Córr.” (surely an abbreviation for córrego) Areio, in the
vicinity of Vila Nova, east of Cuiabá, Mato Grosso State, Brazil. According to Knaack
(2000: 47–48), C. areio seems to be found in two streams that cross the road MT-373,
one of them in the kilometer 8.5 and the other one in the kilometer 13.5, near the city
of Poxoréo. Searches for “Vila Nova” in Mato Grosso only resulted in a small town at
the margins of the rio das Garças, upper rio Araguaia basin, which seems unlikely to be
related to the C. areio type-locality. Even though it was not possible to track these exact
points, the córrego Areia basin, tributary from the rio Poxoréo, itself a tributary from
rio Vermelho, where the rio São Lourenço flows seems the most plausible potential area
for the type-locality, especially considering that the córrego Areia itself and some of
its tributaries cross the road MT-373 close to the city of Poxoréo (in a range of about
10 kilometers). In a recent collecting trip led by LFCT, the córrego Areia was sampled
where it crosses the road MT-130, less than 1 km from the urban area of Poxoréo, where
no specimens of C. areio were captured. Despite this, considering the data provided by Knaack (2000), it seems reasonable to consider the córrego Areia drainage around Poxoréo (15°50’41”S 54°26’44”W) as the type-locality of C. areio .
Another curious subject regarding the C. areio original description refers to the institution where the holotype and nine paratypes were deposited, which was not mentioned in detail, having the catalogue number as the only available information, BZM 33113 and BZM 33136 , respectively. A search in the databases available in Sabaj (2019) and Fricke, Eschmeyer (2020) revealed no match for the acronym “ BZM ”. However, Knaack himself (1966: 364) provided further information on “ BZM ” as follows: “Das Material, welches der Neubeschreibung von Eigenmann und Ward [1907] zu Grunde lag, wurde von Anisits in Paraguay eigesammelt und davon Paratypen auch dem Berliner Zoologischen Museum überlassen (Asuncion, Pilcomayo, BZM Nr. 17249; Matto Grosso, Corumba, BZM Nr. 17258)” (= The material on which the new description by Eigenmann, Ward (in Eigenmann et al., 1907) was based was collected by Anisits in Paraguay, and the paratypes were also given to the Berlin Zoological Museum (Asunción, Pilcomayo, BZM 17249 ; Mato Grosso , Corumbá , BZM 17258 )). Therefore , it seems reasonable to conclude that Knaack created “ BZM ” for “Berliner Zoologischen Museum”, not knowing that the “Zoologischen Museum” in Berlin (a former name of the Museum für Naturkunde ) was recognized by the acronym “ ZMB ” (see Sabaj , 2019) .
Although Knaack (2000:47) planned to send the type series of C. areio to a fish collection in São Paulo (possibly MZUSP): “Es ist vorgesehen den Holotyp und weiteres Typenmaterial dem Museum Sao Paulo zu übergeben” (= It is planned to hand over the holotype and other type material to the São Paulo Museum), the holotype and paratypes were repatriated in 2001 through their deposit at the MCP, in Rio Grande do Sul, Brazil, under the catalog numbers MCP 28675 View Materials and MCP 28676 View Materials , respectively. Additionally, four topotypes were also donated by the ZMB ( MCP 28677 View Materials ) (Carlos A. S. Lucena, 2020, pers. comm.) .
Material examined. All from Brazil, rio Paraguay basin. Mato Grosso: MNRJ 20694 View Materials , 2 View Materials , 27.0– 41.5 mm SL, Dom Aquino , ribeirão Parnaíba, tributary of the rio São Lourenço, 15º44’22”S 54º56’44”W, F. A. G. Melo, P. A. Buckup & M GoogleMaps . R. S. Melo , 18 Feb 2000 . MZUSP 36720 View Materials , 6 View Materials , 27.7–37.1 mm SL, Itiquira , rio Piquiri , Porto Chinelo, Fazenda Santo Antônio do Paraíso, J. H. B. Medeiros & J. C. Oliveira, 17–20 Oct 1980 . MZUSP 75231 View Materials , 2 View Materials , 26.1–36.5 mm SL, Itiquira , rio Piquiri e Itiquira, Fazenda Santo Antônio do Paraíso, J. C. Oliveira & J. H. B. Medeiros. Mato Grosso do Sul : MNRJ 51964 View Materials , 5 View Materials , 34.2–40.1 mm SL, Pedro Gomes , unnamed stream tributary from the córrego do Lobo, rio Taquari basin 17°50’22”S 54°04’19”W, L. F. C. Tencatt, M. N. Souza & M. Santos, 28 Nov 2018 GoogleMaps . MZUSP 59364 View Materials , 3 View Materials , 22.9 View Materials –25.0 mm SL, Aquidauana , rio Taboco, rio Negro basin, 20º04’18”S 55º38’47”W, A. Machado-Allison, B. Chernoff, O. Froehlich & A. Catella GoogleMaps . ZUFMS 1105 , 5 , 33.7–44.4 mm SL, Corguinho , córrego São João (= córrego Periquito), rio Taboco basin, 19°56’11”S 55°19’28”W, O. Froehlich, M GoogleMaps . R. Cavallaro, D. Silva, L. S. Inocêncio & A. Brum, 11 Oct 2001 . ZUFMS 1314 , 15 , 34.4–41.9 mm SL, 2 cs, 38.1-38.7 mm SL, Corguinho , córrego São João (= córrego Periquito), rio Taboco basin, 19°56’11”S 55°19’28”W, O. Froehlich, M GoogleMaps . R. Cavallaro, J. Sedenho, D. Silva, M . V. Costa & F. L.G. Leal, 2 Jun 2001 . ZUFMS 3899 , 3 , 25.8–36.9 mm SL, Corguinho , córrego São João (= córrego Periquito), rio Taboco basin, 19°56’12”S 55°19’27”W, L. F. C. Tencatt, B. F. Santos GoogleMaps & R. L. Santos , 18 Jul 2014 . ZUFMS 4134 , 2 , 27.6–30.7 mm SL, Aquidauana , rio Taboco, rio Negro basin, 20°00’15”S 55°20’53”W, L. F. C. Tencatt, B. F. Santos GoogleMaps & T. R. F. Sinani, 4 Dec 2014 . ZUFMS 4135 , 1 , 34.3 mm SL, Aquidauana , rio Taboco, rio Negro basin, 20°00’15”S 55°20’53”W, L. F. C. Tencatt, B. F. Santos GoogleMaps & T. R. F. Sinani, 19 Jan 2015 . ZUFMS 4136 , 1 , 36.0 mm SL, Aquidauana , córrego Mundo Novo (= córrego Grande), rio Taboco basin, 20°01’45”S 55°15’43”W, L. F. C. Tencatt, B. F. Santos GoogleMaps & T. R. F. Sinani, 4 Dec 2014 . ZUFMS 4137 , 13 , 30.2– 40.8 mm SL, Corguinho , córrego Bandeira, rio Taboco basin, 19°51’45”S 55°16’16”W, L. F. C. Tencatt, B. F. Santos, F. Severo-Neto GoogleMaps & T. R. F. Sinani, 20 Apr 2015 . ZUFMS 5951 , 7 , 30.3–42.3 mm SL, Corguinho , córrego Bandeira, rio Taboco basin, 19°51’45”S 55°16’16”W, L. F. C. Tencatt, B. F. Santos, F. Severo-Neto GoogleMaps & T. R. F. Sinani, 20 Apr 2015 . ZUFMS 5965 , 1 , 36.9 mm SL, Corguinho , córrego São João (= córrego Periquito), rio Taboco basin, 19°56’12”S 55°19’27”W, L. F. C. Tencatt, B. F. Santos GoogleMaps & R. L. Santos , 18 Jul 2014 . ZUFMS 6379 , 3 , 37.7–40.8 mm SL, Pedro Gomes , unnamed stream, rio Taquari basin, 17°57’39”S 54°10’39”W, L. F. C. Tencatt, M. N. Souza & M. Santos, 28 Nov 2018 GoogleMaps . ZUFMS 6380 , 1 , 32.4 mm SL, Coxim , ribeirão dos Veados (= córrego do Veado), rio Taquari basin, 18°25’21”S 54°50’04”W, L. F. C. Tencatt, H. Gimênes Jr GoogleMaps ., R. Rech & R. Mochi , 16 May 2018 . ZUFMS 6381 , 5 , 32.4–36.3 mm SL, Pedro Gomes , unnamed stream tributary from the córrego do Lobo, rio Taquari basin 17°50’22”S 54°04’19”W, L. F. C. Tencatt, M. N. Souza & M. Santos, 28 Nov 2018 GoogleMaps .





| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.