
Compsobuthus arabicus Levy, Amitai & Shulov, 1973
|
publication ID |
https://doi.org/ 10.5281/zenodo.5741445 |
|
publication LSID |
lsid:zoobank.org:pub:15B1EA02-BFD2-43CB-80FD-B8B12BA6C0BC |
|
DOI |
https://doi.org/10.5281/zenodo.6546401 |
|
persistent identifier |
https://treatment.plazi.org/id/3133B07B-FFA4-106A-9456-A6CF5916FAB9 |
|
treatment provided by |
Felipe |
|
scientific name |
Compsobuthus arabicus Levy, Amitai & Shulov, 1973 |
| status |
|
Compsobuthus arabicus Levy, Amitai & Shulov, 1973 View in CoL
( Figures 1–63 View Figures 1–9 View Figures 10–17 View Figures 18–22 View Figures 22–37 View Figures 38–54 View Figures 55–64 , 65–109 View Figure 65 View Figure 66 View Figure 67 View Figure 68 View Figures 69–77 View Figures 78–84 View Figures 85–93 View Figures 94–102 View Figures 103–104 View Figures 105–106 View Figures 107–108 View Figure 109 , 216–218 View Figures 210–224 , Tables 1–2 View Table 1 View Table 2 , 4 View Table 4 )
Compsobuthus arabicus Levy, Amitai & Shulov, 1973: 122– 124 View in CoL , figs. 17–20; Fet & Lowe, 2000: 125 (complete reference list until 1998); Tigar & Osborne, 1999: 174, 180, tab. 2; Lowe, 2001: 172; Fet et al., 2003: 3; KovařÍk, 2003: 89 (?); Hendrixson, 2006: 62–64, figs. 10, 13; Lowe, 2009: 3; El-Hennawy, 2009: 121 (in part); Lowe, 2010: 36; Lourenço & Duhem, 2012: 124; KovařÍk, 2012: 2; KovařÍk & Ojanguren-Affilastro, 2013: 146–149 (in part); Alqahtani et al., 2019: 22, fig. 2d (?); Lowe et al., 2019: 24, fig. 108.
Compsobuthus acutecarinatus arabicus: KovařÍk, 2001: 80 View in CoL .
Compsobuthus acutecarinatus: KovařÍk, 2002: 7 View in CoL (in part).
TYPE LOCALITY AND TYPE DEPOSITORY. Saudi Arabia, Daugha ; BMNH.
TYPE MATERIAL. Saudi Arabia, Daugha [18.9833°N 51.1134°E], 10.I. 1931, 900 ft a.s.l., 1♀ (holotype, 1931.6.2.20, examined), leg. B.S. Thomas, BMNH; GoogleMaps Ramlat Mefel [19.9536°N 54.5516°E, 114 m a.s.l.] 1♀ (examined), GoogleMaps Khor Mefel [19.9117°N 54.4814°E, 114 m a.s.l.] 1♀ (not examined), X.1945 – II.1946, leg. W. Thesiger, BMNH. GoogleMaps Oman: Wadi Mughshin [19.5476°N 54.8835°E, 114 m a.s.l.], 1♀ (not examined), X.1945 – II.1946, leg. W. Thesiger, BMNH. GoogleMaps
REMARKS. Locality coordinates of type material cited above (in square brackets) were obtained from maps published by the collectors ( Thomas, 1931; Thesiger, 1946). Two paratype localities of Thesiger cited as ‘Ramlat Enfel’ and ‘Khor Enfel’ in the original description of Levy et al. (1973) were misspelt. In fact, Thesiger (1946) indicated on his map the localities Ramlat Mefel and Khor Mefel along the path of his travels. These two sites are near each other, positioned within the dunes of the Rub’ al-Khali in southeastern Saudi Arabia, close to the Oman border. Errors may have been due to misreading of labels, and were duplicated in subsequent listings ( Fet & Lowe, 2000; El-Hennawy, 1992, 2009; Vachon, 1979; Alqahtani et al. 2019). The other paratype locality cited by Levy et al. (1973), ‘Wadi Mughhin’, is actually Wadi Mughshin ( Thesiger, 1946) (also spelt ‘Wadi Muqshin’), as suggested by Fet & Lowe (2000). KovařÍk (2003) listed C. arabicus (1 ♂, immature) from 150 km SSW of Riyadh, central Saudi Arabia (see also KovařÍk, 2002: 7). We have not re-examined this specimen to confirm its validity. Alqahtani et al. (2019) reported C. arabicus from southwestern Saudi Arabia, although we have not directly confirmed this record.
OTHER MATERIAL EXAMINED. Oman: Mintirib, 22°26'N 58°48'E, 268 m a.s.l., 13.II.1980, wadi, Oman Eastern Sands Project, 1♀, leg. W. Büttiker, NHMB 705.00; GoogleMaps Wahiba Sands, 21°57'N 58°53'E, 22.XII.1985, 1♀, leg. M. D. Gallagher, NHMB; GoogleMaps Shariq, 22°28'N 58°48'E, 295 m a.s.l., 9.II.1986, Acacia wood, Oman Eastern Sands Project, 1♀, leg. W. Büttiker, NHMB; GoogleMaps Yalooni , 19°56'N 56°06'E, 2.XI.1993, 1♀, leg. Cjv, YAL 518, BMNH GoogleMaps ; Yalooni, 19°57'N 57°07'E, 154 m a.s.l., 18.XI.1993, inside portacabin, 1♂, leg. M. W. Lawrence, NHMB; GoogleMaps Wahiba Sands, 22°10.68'N 58°51.16'E, 90 m a.s.l., 7.X.1993, 19:00 h, UV detection, base of shrub, nr base of linear dunes with vegetation, 2♂, leg. G. Lowe, M.D. Gallagher, N. Wood and S. Prakash, NHMB; GoogleMaps Yalooni, Jiddat Al Harasis , 19°56'N 57°05'E, 154 m a.s.l., 14.III.1994, 22:00 h, on sand and stones, 1♀, leg. M.D. Gallagher, MDG 8561, GLPC GoogleMaps ; Yalooni, 19°56'N 57°05'E, 154 m a.s.l., 15.III.1994, 22:00–23:00 h, UV detection, still on plant remains, under Acacia bush on sand mound, clear sky, new moon, slight breeze, 1♀, leg. M. D. Gallagher, MDG 8564.5, NHMB; GoogleMaps W of Ghabah , 21°23.89'N 57°09.56'E, 185 m a.s.l., 5.X.1994, UV detection on sand, sandy wadi with dunes and scrub adjacent to sabkha, windy, 2♂, leg. G. Lowe and M. D. Gallagher, MCZ GoogleMaps ; NW of Ghabah , 21°31.55'N 57°16.49'E, 180 m a.s.l., 5.X.1994, UV GoogleMaps detection in small wadi, flat gravel plain, 1♂ 1♀, leg. G. Lowe and M. D. Gallagher, ONHM ; Ramlat As Sahmah, 20°06.6'N 55°58.89'E, 155 m a.s.l., 7.X.1994, UV detection on sand, low dune slope, vegetated hummocks with Calligonum , 1♂, leg. G. Lowe and M. D. Gallagher, NHMB; GoogleMaps S of Ghabah , 21°20.99'N 57°14.52'E, 180 m a.s.l., 8.X.1994, UV detection in sandy wadi with bushes, 1♂, leg. G. Lowe and M. D. Gallagher, GLPC GoogleMaps ; between Ghabah & Adam , 22°09.54'N 57°30.16'E, 250 m a.s.l., 16.IX.1995, 22:00-23:00 h, UV detection, sandy vegetated wadi in open gravel plain, trees and bushes, 1♂, leg. G. Lowe and J. Dundon, ONHM GoogleMaps ; nr Wadi Andam, road south of Sinaw , 21°19.35'N 58°15.55'E, 90 m a.s.l., 19.IX.1995, 23:00 h, UV detection on soft sand at base of shrubs, sandy soil mixed with firmer soil, gravel and small stones, patch of vegetation with shrubs and small trees, scorpions run quickly on sand when exposed to UV light, 1♀, leg. G. Lowe and M. D. Gallagher, NHMB; GoogleMaps nr Wadi Andam, road south of Sinaw , 21°19.48'N 58°15.24'E, 90 m a.s.l., 19.IX.1995, 23:30 h, UV detection on soft sand at base of shrubs, sandy soil mixed with firmer soil, gravel and small stones, patch of vegetation with shrubs and small trees, 1♂, leg. G. Lowe and M. D. Gallagher, NHMB; GoogleMaps 10 km N Adam , 22°29.66'N 57°33'E, 300 m a.s.l., 23.X.1995, 22:45 h, in the open on sandy or gritty ground in a well vegetated small wadi, 1♀, leg. J. Dundon, GLPC GoogleMaps ; 55 km NW Ibri , 23°36.5'N 56°05.33'E, 290 m a.s.l., 22.XI.1995, 19:30-20:30 h, area of low sand dunes, scorpions found at entrance to burrows, 2♂, leg. J. Dundon 106, NMPC GoogleMaps ; 55 km NW Ibri, 23°35.5'N 56°04.33'E, 290 m a.s.l. 22.XI.1995, 20:30–21:30 h, area of low sand dunes, scorpions found at entrance to burrows, 1♂, leg. J. Dundon 107, NHMB; GoogleMaps between Qarn Alan and Ghabah North , 21°22.03'N 57°05.47'E, 150 m a.s.l., 21.II.1996, UV detection on coarse grit on top of dusty alluvium, in shallow depression, Acacia ehrenbergiana with sand mounds at base, 1♂ 2♀ 1 juv., leg. M. D. Gallagher, MDG 8755 , NHMB; GoogleMaps 30 km S. of Adam, 22°05.9'N 57°31.12'E, 19.III.1996, 23:30-01:00 h, in sandy wadi, 2♀, leg. J. Dundon 118, NHMB; GoogleMaps Ghneem (= Saiwan ), 20°53.75'N 57°38.78'E, 17.IV.1997, 19:00-22:00 h, open sand between gravel ridges of small hills, 1♂, leg. M. D. Gallagher, I. D& M. Harrison and J. Peterson, MDG 8857, NHMB; GoogleMaps Yalooni, 19°56'N 57°06'E, 1.VII.1997, Yalooni camp, 1♀, leg. S. Brend, YAL 617 , NHMB; GoogleMaps Ramlat Muqshin, 19°46.3'N 55°07.1'E, 100 m a.s.l., 29.XI.1997, on small dune sand, most under cover of scrub bushes, new moon, 4♀, leg. M. D. Gallagher and I. D. Harrison, MDG 8905 , NHMB; GoogleMaps Al Mushash , 19°39.5'N 54°00'E, 129 m a.s.l., 1.XII.1997, under rubbish on sand, 1♀, leg. M. D. Gallagher, MDG 8910, USNM GoogleMaps ; Wadi Qitbit , 19°07'N 54°31'E, 100 m a.s.l., 6.XII.1997, dry sandy desert with shrubs and small trees, strong new moon, no breeze, 1♀, leg. I. D. Harrison and M. D. Gallagher, MDG 8923, ONHM GoogleMaps ; 15 km NNE of Fasad , 18°45.2'N 53°08.9'E, 290 m a.s.l., 29-30.I.1998, in high dunes and lower slopes, 1♂ 1♀, leg. M. D. Gallagher and J. N. Barnes, MDG 8940, GLPC GoogleMaps ; N. of Fasad, Empty Quarter , 18°40.3'N 53°04.4'E, 31.I.1998, under stone on level sand between dunes, 1♀, leg. J. N. Barnes, NHMB; GoogleMaps E. of Ghabah Rest House , 21°22.8'N 57°15.2'E, 130 m a.s.l., 3.I.1999, 17:45 h, rocky plain with coarse grit, under rock, 1♀, leg. A. Winkler, ZSMC GoogleMaps ; Wadi Qit Bit , 19°08.11'N 54°31.08'E, 212 m a.s.l., 18.XII.2001, 16:30-17:45 h, day collection, under oil barrel, 2♀, leg. A. Winkler, ZSMC GoogleMaps ; Wadi Qit Bit , 19°09.33'N 54°30.47'E, 210 m a.s.l., 18.XII.2001, 20:00-21:00 h, UV detection, sandy dunes, near spring, on sand between shrubs, 1♂, leg. A. Winkler, ZSMC GoogleMaps ; W of Wahiba sands, 22.02990°N 58.18490°E, IX.2016, 1♂ (captive bred, 1801), 1♀, leg. M. Stockmann, FKCP GoogleMaps ; Al Wasil , 22.49143°N 58.71513°E ( Fig. 107 View Figures 107–108 ), XI.2017, dry dune area, sandy vegetated areas, UV detection at base of small shrubs, 1♂ (captive bred), 2♀, leg. M. Stockmann, FKCP GoogleMaps ; SW of Wahiba, 21°19.35'N 58°15.55'E, XI.2017, nr Wadi Andam , sandy area with small shrubs, 2♀, leg. M. Stockmann, FKCP GoogleMaps ; Dhofar Province, 236 km N Salalah, Wadi Qitbit , 19°9'20"N 54°30'27"E, 160 m a.s.l., 16-17.I.2018, 1♀, leg. P. KabÁtek, FKCP GoogleMaps ; E of Aydam , 17.888736°N 53.066401°E ( Fig. 108 View Figures 107–108 ), X.2019, 1♂ (No. 1789), leg. M. Stockmann. GoogleMaps United Arab Emirates: Bada Zaid , Abu Dhabi, 24°15'N 54°28'E, 20. V.1972, 1♀, leg. D. J. G. Williams, RS 6512, MNHN GoogleMaps ; Madinet Zayed , 23.68167°N 53.69861°E, 14.X.1993, 1♂ 1♀, leg. A. Saji, TERC GoogleMaps ; Al Khatim , 24.16849°N 54.98412°E, 17.XI.1993, 1♂, leg. A. Saji, TERC GoogleMaps ; env. Lahhab , Dubai, 24°59'00.7"N 55°39'36.2"E, 151 m a.s.l., 21.XI.2006, 1♂, leg. J. Batelka and H. Pinda, FKCP GoogleMaps .
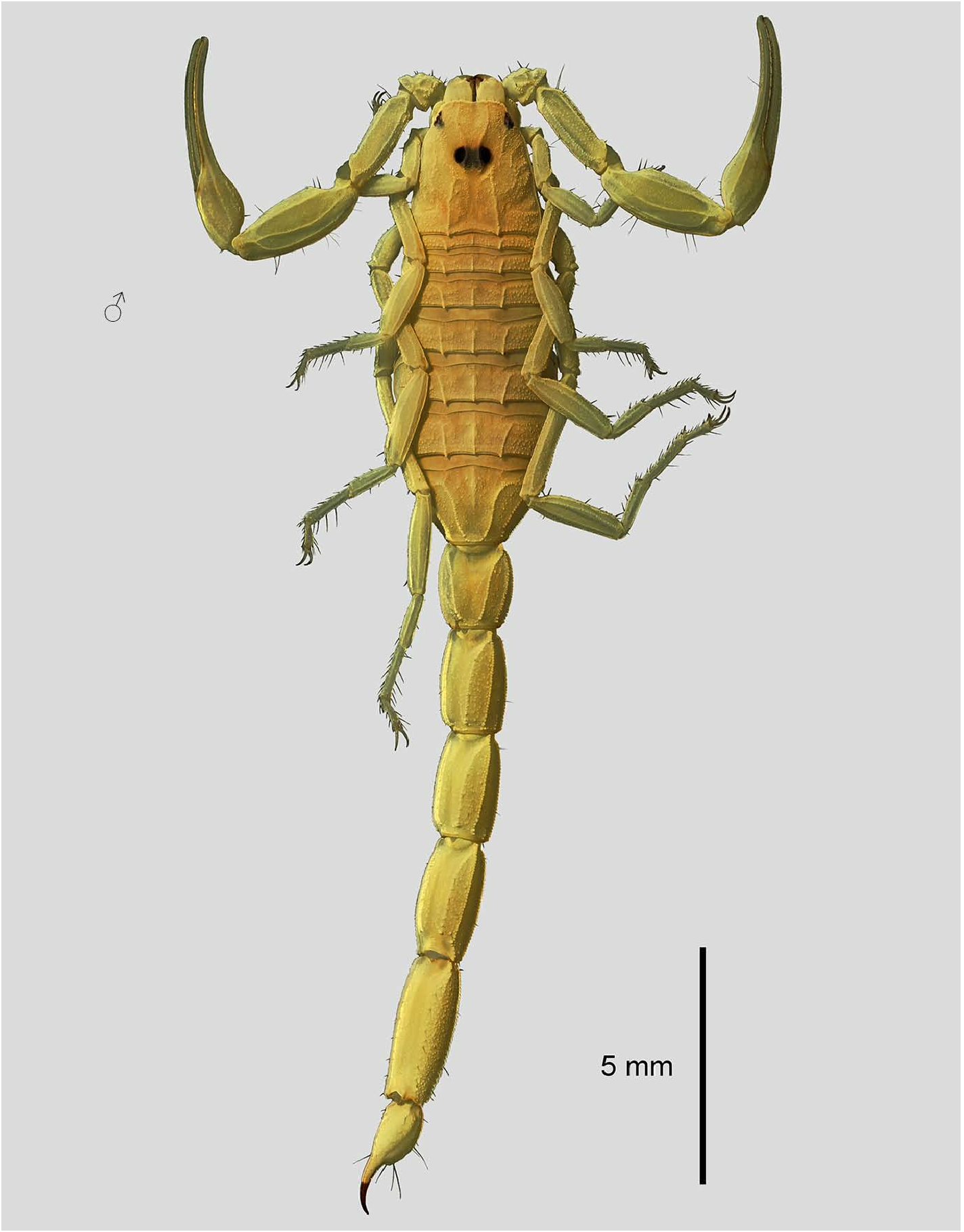
DIAGNOSIS. Total length 20–29 mm. Sexual dimorphism minor, pedipalp fingers straight in females, almost straight or weakly scalloped in males, male chela with broader manus, chela L/W ratio: ♂ 4.70–5.47, ♀ 5.61–6.55; metasomal segment proportions similar in both sexes. Base color uniform yellow. Carapace, tergites, pedipalps and legs densely, finely granular or shagreened. Anterior margin of carapace bearing 8–10 symmetrically distributed spinules. Pedipalp femur L/ Carapace L ratio: 0.76–0.87. Movable finger of pedipalp chela with 7–8 rows of granules, without external accessory denticles, with 9 internal accessory granules (‘ acutecarinatus ’ group of Levy & Amitai, 1980). Manus of pedipalp chela shorter than fixed finger. Pedipalp chela L/movable finger L ratio: ♂ 1.37– 1.47, ♀ 1.28–1.41. Metasoma I with 10 carinae, II–IV with 8 carinae. Median lateral carinae of segment II replaced by isolated granules that may coalesce into carinae posteriorly. All metasomal segments longer than wide; metasoma L/W ratios: III 1.63–1.82, IV 1.93–2.19, V 2.37–2.80. Metasoma V W/D ratio: 1.07–1.17. Ventral intercarinal surfaces of metasoma lacking macrosetae. Pectine teeth: ♂ 13–16, ♀ 9–15. Pectine L/ Metasoma V W ratio: ♂ 1.40–1.88, ♀ 1.38–1.69. Sternites and metasoma granulated. Sternite VI with 4 weak carinae, VII with 4 crenulate carinae. Telson elongate, aculeus shorter than vesicle. Subaculear tubercle moderate.
DESCRIPTION. Total length 21–29 mm in both sexes. The habitus is shown in Figs. 1–2 View Figures 1–9 , 18–21 View Figures 18–22 , 65–68 View Figure 65 View Figure 66 View Figure 67 View Figure 68 , 103–104 View Figures 103–104 . Trichobothriotaxy of pedipalps is shown in Figs. 10–14, 16 View Figures 10–17 , 39–42, 44–45 View Figures 38–54 , 78–82 View Figures 78–84 .
Sexual dimorphism. Sex differences are minor, females with straight pedipalp fingers ( Figs. 11 View Figures 10–17 , 48 View Figures 38–54 ), male fingers nearly straight, weakly undulate proximally with small gap ( Figs. 39 View Figures 38–54 , 81 View Figures 78–84 ). No sex differences in proportions of metasomal segments. Females with shorter pectines, smaller pectine teeth, larger genital opercula.
Coloration ( Figs. 1–68 View Figures 1–9 View Figures 10–17 View Figures 18–22 View Figures 22–37 View Figures 38–54 View Figures 55–64 View Figure 65 View Figure 66 View Figure 67 View Figure 68 ). Base color is uniform yellow. Variable dark pigmentation may occur on the interocular triangle of carapace, pedipalp femur and patella, legs, and metasomal carinae. Metasoma V can have weak fuscosity that is barely visible.
Carapace and mesosoma ( Figs. 7–8 View Figures 1–9 , 22–27 View Figures 18–22 View Figures 22–37 ). The entire surface of the carapace is densely covered by granules of different sizes.The carinae are moderately to strongly developed and granular. The anterior margin of the carapace is weakly concave medially, and bears 8–10 symmetrically distributed spinules (macrosetae). The tergites are strongly granulated. Tergites I–VI are tricarinate, with strong, denticulate median and lateral carinae. Each carina terminates in a spiniform process that in the lateral carinae extends well past the posterior margin of the tergite. Tergite VII is pentacarinate, with lateral pairs of carinae strong, serratocrenulate, median pairs moderate, crenulate; median carina is weak and confined to the anterior half of the segment. Pectinal tooth counts: ♂ 13–16 (8×13, 9×14, 12×15, 4×16; N = 33 combs), ♀ 10–15 (1×10, 2×11, 17×12, 17×13, 14×14, 1×15; N = 52 combs). The pectine marginal tips extend barely to the posterior margin of sternite III in females, and to half the length of sternite IV in males (not past the distal end of coxa IV). The pectines have 3 marginal lamellae and 6–8 middle lamellae. The lamellae bear numerous dark setae, and each fulcrum bears 2–3 dark setae. Sternites are finely granulated or shagreened, more strongly so on lateral areas of sternites III–VI which have smoother medial areas, and uniformly granulated on sternite VII. The posterior areas of sternites lack a broad glabrous patch. Sternites VI–VII bear 4 crenulate carinae, weakly developed on VI and well developed on VII. Other sternites bear one pair of weak posterior carinae on the medial side of the spiracles.
Metasoma and telson ( Figs. 3–5 View Figures 1–9 , 55–63 View Figures 55–64 ). Metasomal segment I with 10 carinae, II–IV with 8 carinae, and V with five carinae. Median lateral carinae of metasoma II are indicated by isolated granules that may coalesce into carinae posteriorly. All segments sparsely setose and densely granulate. Accessory rows of granules may be present on dorsal surfaces of segments as well as on the ventral surface of segment V. The telson is elongate, with aculeus slightly shorter than vesicle, and a moderately developed subaculear tubercle.
Pedipalps ( Figs. 10–17 View Figures 10–17 , 38–54 View Figures 38–54 ). The pedipalps are finely granulated and sparsely hirsute. The femur bears 5 carinae, the patella 7 granular carinae, the chela 7 carinae. The movable and fixed fingers bear 7–9 rows of granules, without external accessory granules, with 9 internal accessory granules on both fingers. Pedipalp chela L/W ratio: ♂ 4.70–5.47, ♀ 5.61–6.55. Manus of chela shorter than fixed finger. Pedipalp chela L/ movable finger L ratio: ♂ 1.37–1.47, ♀ 1.28–1.41.
Legs ( Figs. 9 View Figures 1–9 , 30–37 View Figures 22–37 ). Legs III–IV bear small tibial spurs. Retrolateral and prolateral pedal spurs are present on all legs. The tarsomeres bear two rows of macrosetae on the ventral surface and several macrosetae on the other surfaces. Sparse bristlecombs of 3–5 setae are developed on basitarsi of legs I–III. The femur bears 4 carinae, the patella 4–6 carinae. The femur and patella bear only solitary macrosetae and are granulated on prolateral surfaces, weakly granulated or smooth on retrolateral surfaces. Tarsal ungues strongly elongated, curved.
Hemispermatophore ( Figs. 72–73 View Figures 69–77 ). Flagelliform, elongate and slender. Flagellum separated from external lobe. Capsule region with 4 lobes at base of flagellum. Posterior lobe longest, triangular, apically rounded, median lobe shortest, apically truncate, anterior lobe acuminate with long thin terminus. Basal lobe is strong with a broad base and sharp, falcate hook. Measurements. Table 1 View Table 1 lists measurements for representative male and female specimens. Morphometric variation is summarized in Table 2 View Table 2 and is plotted in Figs 85–102 View Figures 85–93 View Figures 94–102 . The plots indicate that two male samples taken from populations in the United Arab Emirates are not morphometrically separable from Oman populations ( Figs. 85–93 View Figures 85–93 ). They also indicate that the female holotype from Saudi Arabia is not morphometrically separable from Oman populations ( Figs. 94–102 View Figures 94–102 ). We therefore consider all of these populations to represent a single species exhibiting continuous variation across its geographic range ( Fig. 109 View Figure 109 ).
AFFINITIES. C. arabicus is one of the smaller members of the ‘ acutecarinatus ’ group (Levy & Amitai, 1980), and it has a correspondingly low pectinal tooth count (10–15 in females). Other Compsobuthus species from Oman in the ‘ acutecarinatus ’ group have higher numbers of pectine teeth. In females, C. acutecarinatus (Simon, 1882) has 20–27 teeth, C. maindroni (Kraepelin, 1901) has 19–21, C. nematodactylus Lowe, 2009 has 28–33 and C. polisi Lowe, 2001 has 16–20. In Oman, C. polisi is a small species (adults 22–35 mm) that is outwardly similar to C. arabicus . It can be differentiated by the presence of numerous short, macrosetae on ventral intercarinal surfaces of the metasoma. It differs morphometrically in having somewhat more elongated (higher L/W ratio) metasomal segments III and IV ( Figs. 86 View Figures 85–93 , 95 View Figures 94–102 ) and pedipalp femur and patella ( Figs. 91 View Figures 85–93 , 100 View Figures 94–102 ), although there is overlap in some of the ratios. Metasomal segments of C. polisi tend to be deeper than those of C. arabicus , as indicated by lower L/D ratios ( Figs. 87, 89 View Figures 85–93 , 96 View Figures 94–102 ) and W/D ratio ( Figs. 93 View Figures 85–93 , 102 View Figures 94–102 ) (see Fig. 64 View Figures 55–64 vs. Figs. 55–63 View Figures 55–64 ). In males, C. polisi is separable from C. arabicus by having pedipalp chelae more slender with longer fingers ( Fig. 92 View Figures 85–93 ). Geographically, C. polisi is distributed more along the coastal regions, whereas C. arabicus is ranges further inland in major aeolian dunes (Wahiba/ Sharqiya Sands, Rub’ al- Khali). Two other similarly small species belonging to the ‘ acutecarinatus ’ group are C. pallidus Hendrixson, 2006 , and C. setosus Hendrixson, 2006 , from northern and central Saudi Arabia, respectively. The former can be differentiated by having 10–11 granule rows on the pedipalp fingers, and the latter by the presence of numerous ventral intercarinal setae on the metasoma and sternite VII ( Hendrixson, 2006).
ECOLOGY ( Figs. 107–109 View Figures 107–108 View Figure 109 ). C. arabicus is a psammophilic species found in the expansive aeolian dune systems of Ramlat al-Wahiba and Rub’ al-Khali. It is not an ultrapsammophile, as it is also distributed over other interior regions of Oman where sand dunes intermingle with silty, gravelly or stony substrates, such as the plateau of Jiddat al Harasis ( Fig. 109 View Figure 109 ). Like other arenicolous desert scorpions, it excavates burrows near the bases of vegetation and emerges at night to take ambush positions near burrow entrances. Tigar & Osborne (1999) examined the relationship between the surface activity of a diverse assemblage of nocturnal arthropods and the lunar cycle in an area near Abu Dhabi. Analysis of data pooled from all surveyed taxa revealed significantly more pitfall trap captures on a new moon, than on a full moon. For C. arabicus , there were 2 records on new moon, and none on full moon. However, this sample is too small to test the hypothesis of moon avoidance for this one species. We have observed surface activity of C. arabicus on both moonlit and dark nights.Among 16 independent records of nocturnal surface activity, 12 nights (75%) were 5 days or less from a new moon. This is consistent with the hypothesis, but is not conclusive because there is likely to be an inherent sampling bias due to collectors preferring to search for scorpions by UV detection on darker nights. Other scorpions that were observed together with C. arabicus in sandy habitats include: Androctonus crassicauda (Olivier, 1807) , Apistobuthus pterygocercus Finnegan, 1932 , Buthacus nigroaculeatus Levy, Amitai & Shulov, 1973 , Picobuthus dundoni Lowe, 2010 , P. wahibaensis Lowe, 2010 , Vachoniolus gallagheri Lowe, 2010 and V. globimanus Levy, Amitai & Shulov, 1973 .
LIFE HISTORY ( Figs. 105–106 View Figures 105–106 ). The following data were obtained from captive rearing in a temperate climate. In a gravid female collected in IX–XI.2016, parturition was recorded in VII.2017, with litter sizes of 7 juveniles. Only two individuals were reared to maturity, at sixth (♂) and seventh (♀) instars. In two gravid females collected in XI.2017, parturition was recorded in VI.2018, with litter sizes of 7 and 10 juveniles. These were reared to maturity by VI–VIII.2018 and IX–XI.2018, at fifth (♂) and sixth (♀) instars.

















| NHMB |
Natural History Museum Bucharest |
| UV |
Departamento de Biologia de la Universidad del Valle |
| MCZ |
Museum of Comparative Zoology |
| NMPC |
National Museum Prague |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ZSMC |
Zoologische Staatssammlung |
| V |
Royal British Columbia Museum - Herbarium |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Compsobuthus arabicus Levy, Amitai & Shulov, 1973
| Kovařík, František, Lowe, Graeme, Stockmann, Mark & Šťáhlavský, František 2020 |
Compsobuthus acutecarinatus: KovařÍk, 2002: 7
| KOVARIK, F. 2002: 7 |
Compsobuthus acutecarinatus arabicus: KovařÍk, 2001: 80
| KOVARIK 2001: 80 |
Compsobuthus arabicus
| ALQAHTANI & ELGAMMAL & GHALEB & BADRY 2019: 22 |
| LOWE, G. & F. KOVARIK & M. STOCKMANN & F. STAHLAVSKY 2019: 24 |
| KOVARIK, F. & A. A. OJANGUREN-AFFILASTRO 2013: 146-149 |
| LOURENCO W. R. & DUHEM 2012: 124 |
| KOVARIK 2012: 2 |
| LOWE 2010: 36 |
| LOWE 2009: 3 |
| EL-HENNAWY, H. K. 2009: 121 |
| HENDRIXSON 2006: 62 |
| FET 2003: 3 |
| KOVARIK 2003: 89 |
| LOWE 2001: 172 |
| FET, V. & G. LOWE 2000: 125 |
| TIGAR, B. J. & P. E. OSBORNE 1999: 174 |
| Levy, Amitai & Shulov 1973: 122-124 |