
Notoglanidium pembetadi, Vreven, Emmanuel, Zamba, Armel Ibala, Mamonekene, Victor & Geerinckx, Tom, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3691.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:DED26CDC-E602-478F-9DFE-7273A264AA8A |
|
DOI |
https://doi.org/10.5281/zenodo.5625943 |
|
persistent identifier |
https://treatment.plazi.org/id/03908789-FFCD-FFAF-FF2A-FC1AFD9F9DC6 |
|
treatment provided by |
Plazi |
|
scientific name |
Notoglanidium pembetadi |
| status |
sp. nov. |
Notoglanidium pembetadi View in CoL new species
Figures 1 View FIGURE 1 & 2 View FIGURE 2 ; Tables 1 View TABLE 1 & 2
Anaspidoglanis boutchangai (in part) non (Thys van den Audenaerde, 1965): Mbega & Teugels (1998); Geerinckx et al. (2003, 2007).
Holotype. MRAC A7-31-P-0029, 150.0 mm SL female, Republic of the Congo, Niari (Kouilou) River at Pont Niari, just downstream of the bridge [i.e., bridge on the road Dolisie-Nyanga], 3° 33.57’ S, 12° 20.47’ E, 0 8 Sep. 2007, Vreven and Ibala Zamba.
Paratypes. Republic of the Congo: AMNH 243475 (formerly MRAC A7-13-P-45–46), 165.7–179.4 mm SL males, same data as for holotype. MRAC A7-13-P- 16–21, 160.8 mm SL female, 53.0– 169.8 mm SL males, Pont du Niari, Kouilou-Niari River, at the bridge on the road Dolisie-Nyanga, hook (purchased), night catch, 3° 33.314’ S, 12° 20.583 ’E, 23 Sep. 2006, Vreven and Ibala Zamba. MRAC A7-31-P-0030–0032, 131.9 mm SL female, 115.9–140.1 mm SL males, same data as for holotype. MRAC A7-13-P-42–44, 153.0– 162.2 mm SL females, 123.0 mm SL male, Pont du Niari, downstream of the bridge, Kouilou-Niari Basin, right bank, rotenone, 3° 33.547’ S, 12° 20.341’ E, 23 Sep. 2006, Vreven and Ibala Zamba.
Non-type specimens. Republic of the Congo: MRAC 91-79-P-283, 71.0 mm SL (cleared and stained), Kouilou-Niari River at pont du Niari near Kibangou, 03° 33’ S, 12° 20’E, 0 2 Oct. 1986, Roberts. MRAC 91-79-P- 284, 68.2 mm SL male, same data. MRAC A5-36-P-119, 111.0 mm SL male, Kouilou-Niari Basin, at Pont du Niari, 3° 33.314’ S, 12° 20.583’ E, 18 Aug. 2005 Mamonekene, Vreven and Ibala Zamba. MRAC A7-13-P-22–28, 34.8–133.5 mm, Pont du Niari, Kouilou-Niari Basin, at the bridge on the road Dolisie-Nyanga, hook (purchased), night catch, 3° 33.314’ S, 12° 20.583’ E, 23 Sep. 2006, Vreven and Ibala Zamba. MRAC A7-13-P-29–41, 86.9– 175.6 mm, Pont du Niari, upstream of the bridge, Kouilou-Niari Basin, right bank, rotenone, 3° 33.273’ S, 12° 20.592’ E, 24 Sep. 2006, Vreven and Ibala Zamba. MRAC A7-13-P-47–63, 33.0– 179.7 mm SL, Pont du Niari, downstream of the bridge, Kouilou-Niari Basin, right bank, rotenone, 3° 33.547’ S, 12° 20.341’ E, 23 Sep. 2006, Vreven and Ibala Zamba (one specimen, MRAC A7-13-P-63, cleared and stained). MRAC A7-13-P-64–67, 90.7– 134.8 mm SL, Pont du Niari, just downstream of the bridge on the road Dolisie-Nyanga, Kouilou-Niari Basin, gillnets, night catch, 3° 33.547’ S, 12° 20.341’ E, 23 Sep. 2006, Vreven and Ibala Zamba. MRAC A7-31-P-0033, 101.2 mm SL male, data same as for holotype.
Diagnosis. Notoglanidium pembetadi new species can be distinguished from all other species in Notoglanidium , and Anaspidoglanis as well, by the following unique combination of characters: a premaxillary toothplate of horseshoe-like shape ( Fig. 3 View FIGURE 3 a) and a combined toothplate width of 20.8–26.4% HL [vs. cardiform and narrower, 11.1–15.3% HL in N. pallidum ( Fig. 3 View FIGURE 3 b), 11.9–18.2% HL in N. thomasi , and 13.7–16.1% HL in N. walkeri ; crescent-like ( Fig. 3 View FIGURE 3 c) and wider, 30.1–35.3% HL in A. boutchangai and 33.5–41.7% HL in A. macrostoma ; and rectangular ( Fig. 3 View FIGURE 3 d) but similar in width, 22.0–26.1% HL in A. akiri ].
Notoglanidium pembetadi can be further distinguished from all Notoglanidium species by the following unique combination of measurements: a short head length, 21.1–22.8% SL (vs. longer, 27.8–29.5% SL in N. pallidum , 24.1–27.0% HL in N. thomasi and 25.3–28.9% SL in N. walkeri ); and a short dorsal-fin base length, i.e. 21.0– 24.4% SL (vs. longer, 18.4–22.4 in N. pallidum and 29.3–34.1% SL in N. thomasi ).
Notoglanidium pembetadi can be further distinguished from all Anaspidoglanis species by the following unique combination of characters: a high number of dorsal-fin rays, II, 10–13 (vs. less, i.e., II, 7 in all Anaspidoglanis species); a long dorsal-fin base length, 21.1–24.4% SL (vs. shorter, 13.9–20.1% SL in all Anaspidoglanis species); a short predorsal distance, 30.1–33.5% SL (vs. longer, 36.3–44.4% SL in all Anaspidoglanis species); and a short prepelvic distance, 40.1–45.4% SL (vs. longer, 46.6–58.2% SL in all Anaspidoglanis species).
Description. The morphometric and meristic data are given in Tables 1 View TABLE 1 & 2 respectively. Detailed comparative data have been added for A. boutchangai as two specimens of this new species (MRAC 91-79-P-283–284) have been erroneously attributed to this species by earlier researchers [see Mbega & Teugels (1998); Geerinckx et al. (2003, 2007)].
General body shape dorso-ventrally depressed, almost of the same depth along the entire body length, generally highest at posterior head region and dorsal fin. Body width gradually decreasing towards caudal peduncle, which is laterally compressed.
Head depressed, not as wide as long. Eyes without a free border. Snout blunt and rounded. Lips well developed. Anterior nostrils situated on edge of upper lip. Anterior border of posterior nostrils with a minute, rudimentary, somewhat conical nasal barbel. A very small supraoccipital process and nuchal shield as well, resulting in a large distance between both (i.e., 31.8–45.3% HL).
Maxillary barbels reaching one or two eye diameters beyond posterior edge of eye. External mandibular barbels reaching at least anterior edge of pectoral fin base. Internal mandibular barbels reaching posterior edge of eye or approximately one eye diameter beyond it (all barbel lengths are negatively allometric).
Humeral process of cleithrum small, short and rather blunt. Axillary pore (see Friel & Vigliotta 2006; Wright & Page 2006: Fig. 2 View FIGURE 2 ) present, small and situated just below lower border of humeral process. Its presence seems to be the first record for the genus as well as the family.
Pectoral and pelvic fins almost in same horizontal plane. Dorsal and anal fins low. Leading edge of pectoral fins finely pointed and not reaching anterior base of rounded pelvic fins. Pectoral spines generally serrated on posterior and anterior edge with respectively 3–11 and 2–11 serrations, with both counts increasing with the size of the specimens. Insertion of pelvic fins situated approximately below middle of dorsal fin. Posterior tip of pelvic fins not reaching anal fin, but reaching beyond anus. All fins, and especially dorsal, pelvic and anal fins fleshy. Dorsal and anal fins mostly fleshy towards their base and pelvic fin towards unbranched first ray. Adipose fin very low and fleshy, almost continuous with the fleshy anteriormost part of caudal fin. Anterior origin of adipose fin difficult to locate but situated approximately above posterior third of or halfway anal fin. Caudal fin rounded.
Lateral line midlateral and marked by small, equidistantly situated, white edged pores. Body entirely covered with randomly distributed small white edged pores. Maximum body size 179.7 mm SL (see additional specimens for details).
There are 9–11 branchiostegal rays; with eight articulating with the anterior ceratohyal, and two with the posterior ceratohyal in a cleared and stained specimen (MRAC 91-79-P-283). The vertebral column includes the complex vertebra, including v–4 (see Chardon et al., 2003), the fifth vertebra which is partially fused to it, and 34 to 38 additional vertebrae including 13–15 abdominal vertebrae and 21–24 caudal vertebrae ( Table 2).
Coloration. In preserved specimens, overall body colour dark brown. Ventral side of head and body whitish. Ventral side of body between anus and caudal-fin base light brown. In some specimens the lighter coloured region includes the ventral half of the body. Dorsal and anal fins dark brown or with distal thirds white to transparent. Pectoral and pelvic fins dark brown except for their distal 1/4 or 1/3 that is also white to transparent. Lips and barbels dark brown except for white to transparent distal parts of barbels. There seems to be an overall increase in fin pigmentation with body size.
In live specimens, the overall body colour of some individuals is far more light brown. In addition, based on a single photograph ( Fig. 2 View FIGURE 2 ) of a freshly collected specimen, some specimens present 7 or 8 transverse series of black spots in life. However, most of the small as well as large sized specimens lack these spots, both when freshly collected or preserved. There are only two preserved specimens [MRAC A5-36-P-119 (111.0 mm SL) and MRAC A7-13-P-29-41 (175.6 mm SL)] where these spots are still visible.
holotype all examined specimens N.boutchangai
holotype all examined specimens
min max n median
Dorsal-fin rays II, 7 II, 7 II,7 5 II,7 Pectoral-fin rays I, 8 I, 8 I,8 5 I,8 Pelvic-fin rays i5 i5 i5 5 i5 Anal-fin rays 15 14 16 5 15 Caudal-fin rays [upper half/lower half (total)] 8+7 (15) 8+7 (15) 8+7 (15) 4 8+7 (15) Abdominal vertebrae 13 13 14 4 13–14 Caudal vertebrae 22 22 22 4 22 Total vertebral count (4+1)+35 (4+1)+35 (4+1)+36 4 (4+1)+35 Gill rakers [lower arch/angle/upper arch (total)] 10+1+3 (14) 10/1/2 (13) 10/1/3 (14) 5 10/1/2 (13) Branchiostegal rays 11 10 11 4 10–11 Distribution. The new species is only known from around the type locality in the Kouilou-Niari River, at Pont Niari, and is probably endemic to the Kouilou-Niari River basin ( Fig. 4 View FIGURE 4 ).
Etymology. “ Pembetadi ” refers to the local Kunyi name with “ tadi ” meaning stone. Based on our brief inquiries the meaning of “ pembe ” remains unclear. Although the etymology has not been fully elucidated its Kunyi name seems to refer to the habitat where it lives, i.e., the large rocks and pebbles covering the bottom of the Kouilou-Niari River at Pont du Niari ( Republic of the Congo) ( Fig. 5 View FIGURE 5 ). The name was chosen to acknowledge that many of the new species discovered are already well known by the local inhabitants, and to express gratitude to the many children and fishermen who helped to collect the type series of this new species for science.
Generic assignment. As the new catfish species has eyes without a free border and a very small supraoccipital process and nuchal plate as well, resulting in a large distance between both (i.e., 31.8–45.3% HL). These characters readily discard the genera Auchenoglanis Günther, 1865 and Parauchenoglanis Boulenger, 1911 as possible candidates for generic assignment of the new species, as species attributed to both these genera have eyes with a free border. In addition, in Auchenoglanis supraoocipital process and nuchal plate are in close contact or overlapping (Teugels et al., 1991a; Risch, 2003; Geerinckx et al., 2007) whereas in Parauchenoglanis both are in contact with, or only slightly separated from each other, i.e., in-between distance <13.6% HL. The four remaining nominal auchenoglanidinae genera, Notoglanidium Günther, 1903 (three species; type species N. walkeri Günther, 1903 ), Liauchenoglanis Boulenger, 1916 (one species; junior synonym of Notoglanidium according to Poll (1957) and Risch (1992, 2003); with type species L. maculatus Boulenger, 1916 ), Platyglanis Daget, 1978 (one species; with type species P. depierrei Daget, 1978 ), and Anaspidoglanis Teugels et al., 1991 a (three species; with type species A. macrostoma (Pellegrin, 1909) all share the generic characters highlighted above as observed in the new species.
The new species fits with the original description of Notoglanidium Günther, 1903 , and, apart from the higher number of dorsal fin rays, within that of Anaspidoglanis Teugels et al., 1991 as well. In their original description of Anaspidoglanis Teugels et al. (1991a) extensively compared the new genus with Auchenoglanis and Parauchenoglanis , but unfortunately omitted any comparison to the other genera in the auchenoglanidinae group. This renders the generic discrimination of these genera rather troublesome. Genus definitions and generic assignments of species have been unclear for Liauchenoglanis (junior synonym of Notoglanidium according to Poll (1957), Risch (1992, 2003), and Geerinckx et al. (2007) and Platyglanis (see Daget, 1978; Mo, 1991; Ferraris, 2007) as well. A review of the genera Notoglanidium , Liauchenoglanis , Platyglanis and Anaspidoglanis is presented by Geerinckx et al. (2013).



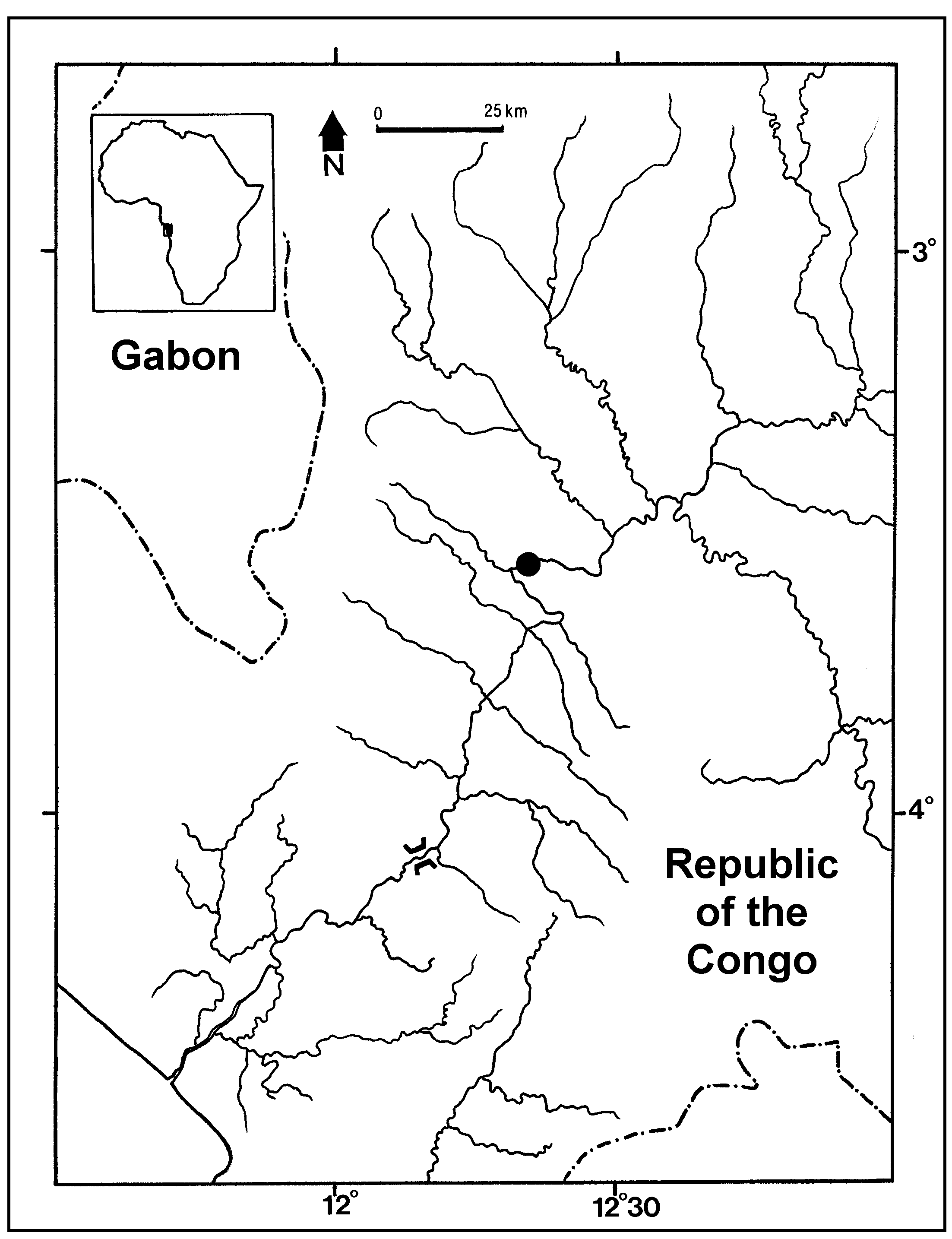

TABLE 1. Morphometric data for the holotype, paratypes and non-type specimens of N. pembetadi and A. boutchangai. Measurements with bold minimum (min), maximum (max) and mean values are diagnostic without overlap of values between both species.
| N. pembetadi | N. boutchangai | |
|---|---|---|
| holotype all examined specimens | holotype all examined specimens | |
| min max n mean | min max n mean | |
| Total length (mm) | 176.0 65.0 212.3 18 149.8 | 238.4 78.8 238.4 5 154.8 |
| Standard length (mm) | 150.0 53.0 179.4 18 127.9 | 194.3 62.3 194.3 5 123.2 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.