
Proteocephalus demshini, Tomáš Scholz & Alain de Chambrier & Takeshi Shimazu & Alexey Ermolenko & Andrea Waeschenbach, 2017
|
publication ID |
https://doi.org/ 10.1016/j.parint.2016.09.0161383-5769 |
|
DOI |
https://doi.org/10.5281/zenodo.6010898 |
|
persistent identifier |
https://treatment.plazi.org/id/632E7F4C-6F79-FF9C-FC9E-05DCFDC6F964 |
|
treatment provided by |
Plazi |
|
scientific name |
Proteocephalus demshini |
| status |
sp. nov. |
3.2.3. Proteocephalus demshini n. sp.
Figs. 4 View Fig. 4 , 6 View Fig. 6
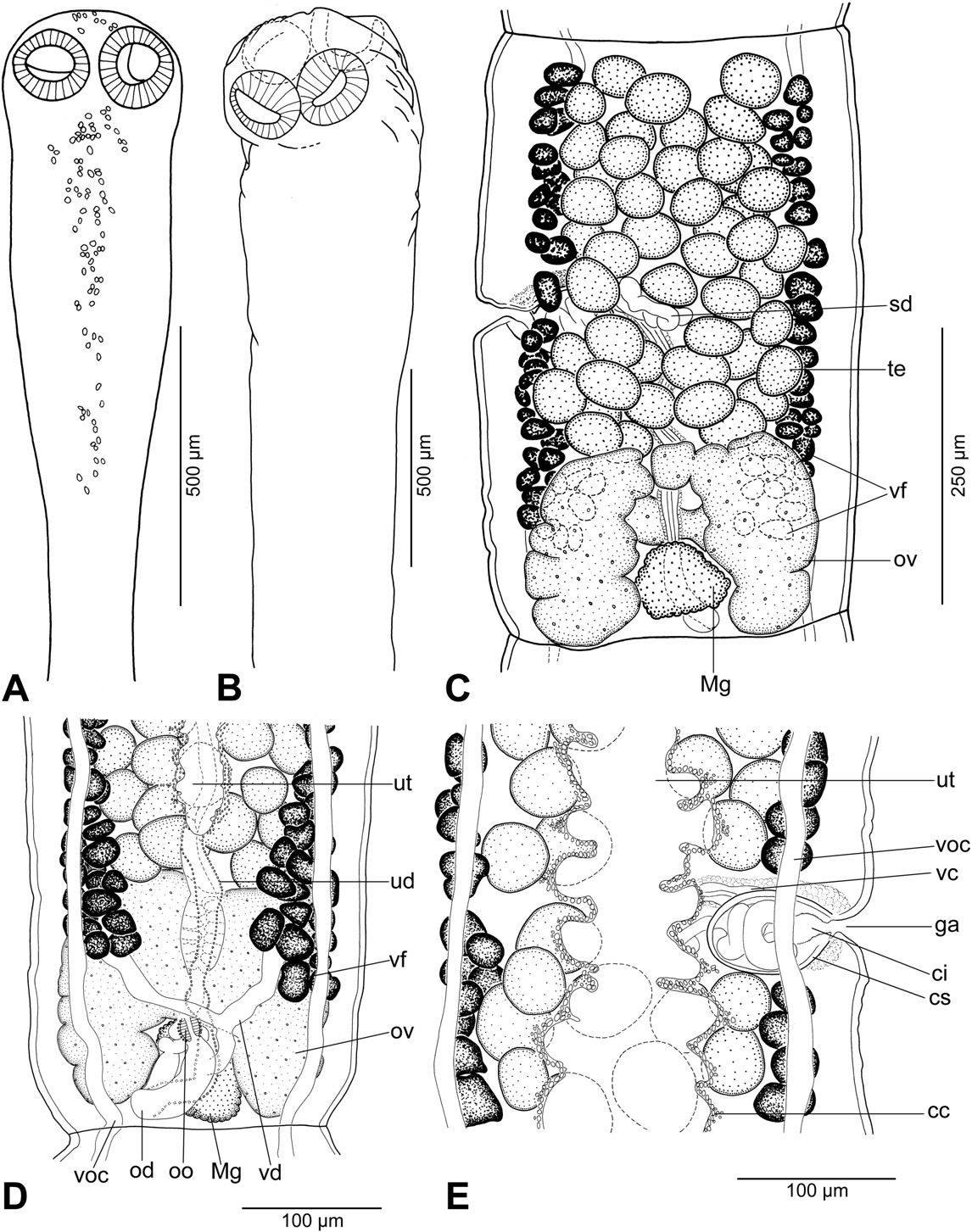
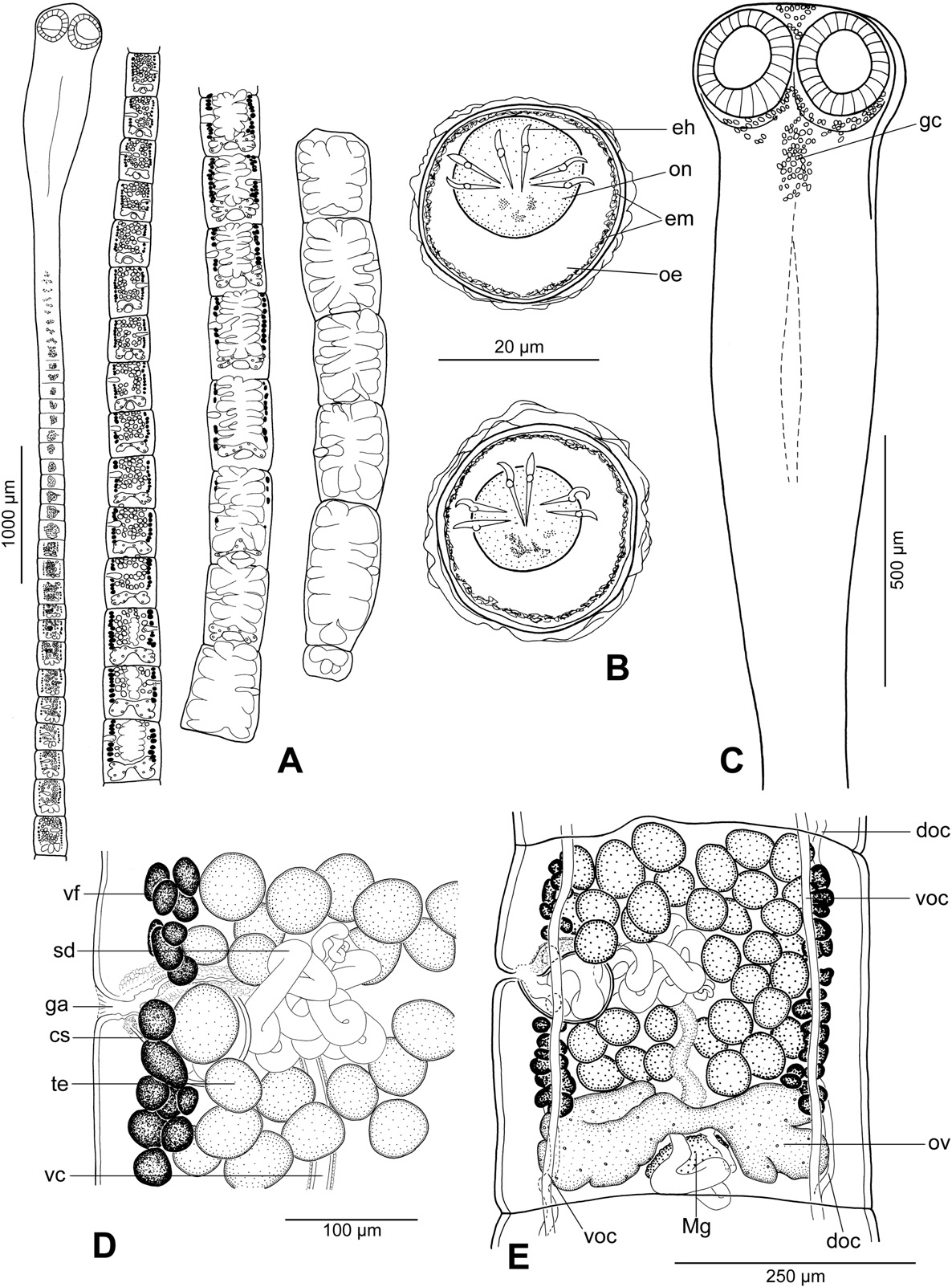
Description (based on 5 whole-mounted specimens and 2 scoleces studied using SEM; for morphometrical data — see Table 2): Proteocephalidae, Proteocephalinae. Testes , vitelline follicles, ovary and uterus medullary. Medium-sized tapeworm, body length 18– 33 mm (n = 5), maximum width up to 1.1 mm (n = 5). Strobila acraspedote, anapolytic, consisting of about 80–100 proglottids: 55–60 immature, 1 mature, 16–24 pregravid and 6–13 gravid proglottids. Immature, mature and pregravid proglottids wider than long, gravid longer than wide.
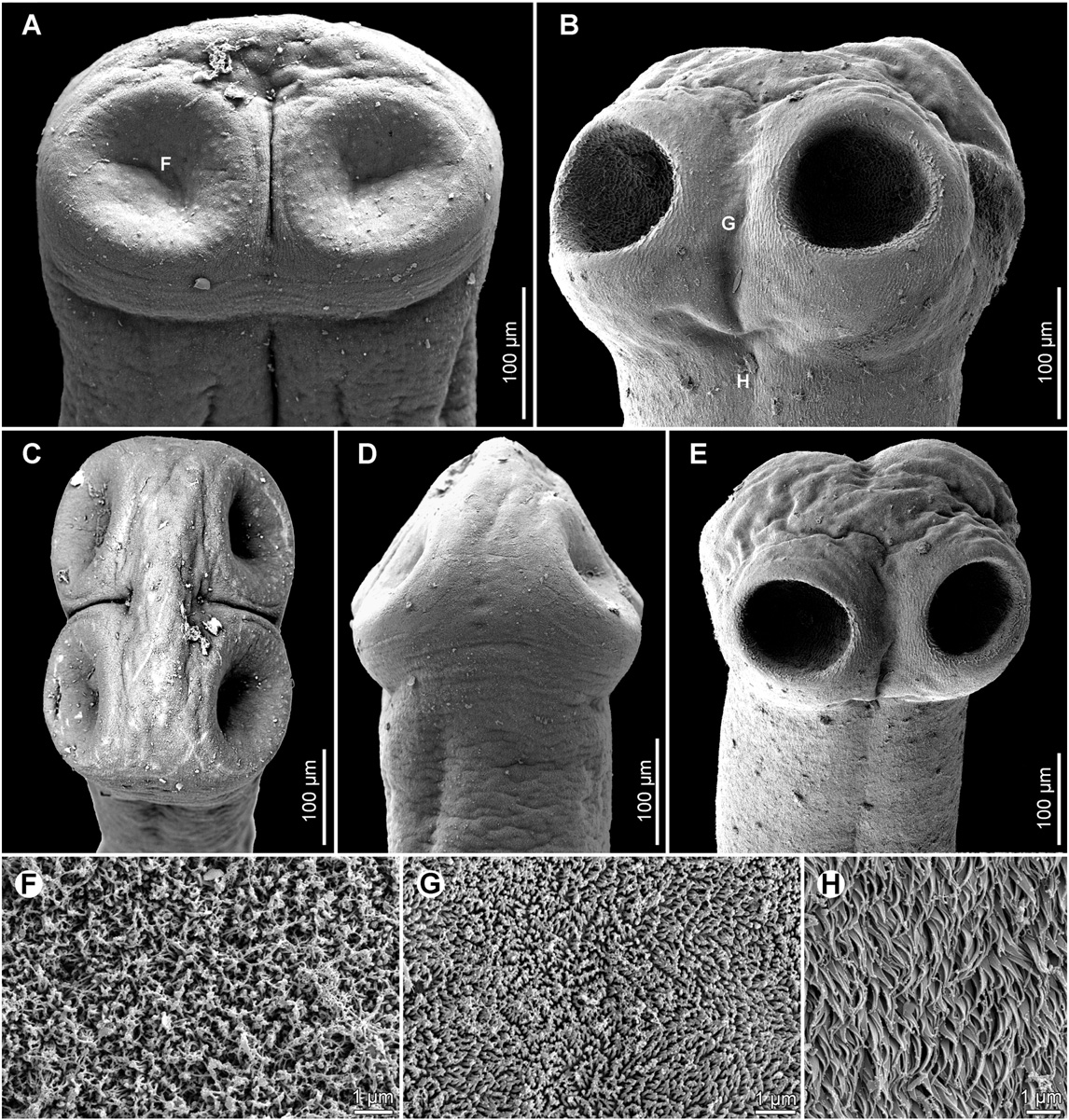
Scolex wider than neck. Suckers spherical, with deep cavity ( Fig.4 View Fig. 4 A).
Apical organ absent; apex of scolex and region posteromedian to suckers with gland cells ( Fig. 4 View Fig. 4 A). Neck (proliferation zone) 360 wide. Scolex covered with acicular filitriches and gladiate spinitriches (on suckers — Fig. 6 View Fig. 6 F) and acicular filitriches (between suckers and on scolex apex); neck covered with gladiate spinitriches.Inner longitudinal internal musculature developed, anastomosed. Ventral osmoregulatory canals thin-walled, 15–25 wide, ventral to lateral bands of vitelline follicles ( Fig. 4 View Fig. 4 C). Dorsal osmoregulatory canals thick-walled, narrow, about 5–8 wide.
Testes 62–89 in number, spherical, in two irregular layers, form one compact field between lateral bands of vitelline follicles and ovary ( Fig. 4 View Fig. 4 B), present also in gravid proglottids. Vas deferens strongly coiled, with loops forming elongated field crossing median line of proglottid ( Fig. 4 View Fig. 4 B). Cirrus-sac pyriform to elongate ( Fig. 4 View Fig. 4 C), thick-walled. Sperm duct (internal vas deferens) coiled. Cirrus muscular. Common genital atrium narrow, deep ( Fig. 4 View Fig. 4 C). Genital pores alternating irregularly, pre-equatorial ( Fig. 4 View Fig. 4 B).
Ovary medullary, bilobed, with long and narrow lateral wings. Relative size of ovary, i.e. its surface in relation to surface of pregravid and gravid proglottids (see [33]), about 17%. Mehlis' gland 85–160 in diameter.
Vaginal canal almost straight in distal part, with terminal part (pars copulatrix vaginae) surrounded by chromophilic cells; vaginal sphincter absent ( Fig. 4 View Fig. 4 C). Vitelline follicles medullary, forming two wide lateral bands interrupted ventrally on poral side at level of cirrus-sac and vagina; bands widened posteriorly and reaching to anterior margin of ovary only, rarely with few follicles slightly (less than by 10%) overlapping ovarian wings on ventral side ( Fig. 4 View Fig. 4 B).
Uterus medullary, with development of type 2 [9]. Lumen of uterine stem occupies up to 49% of proglottid width in last immature proglottids. In mature proglottids, lumen occupies up to 55% of proglottid width; apex lined with numerous chromophilic cells ( Fig. 4 View Fig. 4 C). In gravid proglottids, uterine diverticula occupy up to 90% of proglottid width.
Eggs spherical, with hyaline outer envelope and bilayered embryophore; oncosphere containing embryonic hooks ( Fig. 4 View Fig. 4 D).
Taxonomic summary
Type and known only host: Barbatula toni (Dybowski, 1869)
( Cypriniformes : Nemacheilidae ).
Site of infection: intestine.
Type locality: River Komissarovka near Kamen-Rybolov, Primorsky Region, Russia (44̊57′22″ N, 131̊42′0 0″ E).
Prevalence and intensity of infection (mean with range and number of fi sh examined in parentheses): 19% (n = 32); 1.7 (1–4).
Deposition of specimens: Holotype (one whole-mounted specimen from host field No. RUS 190 collected on 28 June 2011; Coll. No. MHNG-PLAT 79145); three paratypes (whole-mounted specimens from hosts field Nos. RUS 191, 194a and 195 collected on 28 June 2011; IPCAS C-727; MHNG-PLAT 79150); one voucher (wholemounted specimen from host field Nos. RUS 190 collected on 28 June 2011; Coll. No. IPCAS C-727); 2 scoleces from host field No. RUS 192 (MHNG-PLAT 79149) used for SEM observations.
Etymology: The species is named in honour of the late N.I. Demshin, Vladivostok, Russia, for his extraordinary contribution to helminthology in the Russian Far East, especially life-cycles of cestodes.
Differential diagnosis. The new species differs from P. midoriensis and P. misgurni from M. anguillicaudatus (see below) by the shape of the proglottids and the cirrus-sac, width of the lateral bands of the vitelline follicles and their posterior extent (see Remarks on P. midoriensis and compare Figs. 3–5 View Fig. 3 View Fig. 4 View Fig. 5 ). Proteocephalus demshini is strikingly similar to P. sagittus from the congeneric host and a few other loaches in its morphology including the shape of proglottids, which are wider than long except for the last gravid proglottids, wide bands of the vitelline follicles widened posteriorly, and an elongate to pyriform cirrus-sac.
These two species can be distinguished from each other by (i) the number of uterine diverticles on each side, 3–8 in P. demshini versus 9–14 in P. sagittus ; (ii) the ratio of the width of the band of the vitelline follicles at the level of the ovary in relation to the width of the proglottid, which is 15–19% (x = 17%) in the new species versus 8–14% in P. sagittus (x = 10%); (iii) shape of the anterior end of the body, with the neck region widened anteriorly in P. demshini versus of the same width in P.sagittus ; and (iv) the posterior extent of the vitelline follicles, which overlap the ovary only slightly in P. demshini (up to 1/10 of the length of the ovary) versus up to 1/ 5 in P. sagittus ( Table 2).
Discrete species status of these apparent sibling species is supported by molecular data (see Fig. 1 View Fig. 1 and Suppl. Figs. 2 View Fig. 2 and 4 View Fig. 4 ); uncorrected pdistances are 0.5% and 9.9% for lsrDNA and cox1, respectively (Suppl. Table 1).
The type host of this new species, B. toni , occurs in Asia and lives in all rivers flowing to the Arctic and Pacific Oceans between the rivers Ob and Huang He [1]. Its helminth fauna is relatively depauperate [3, 34–36], but includes also three species of cestodes, yet none of them being specific to, or typical of, this fish. Scholz and Ergens (1990) [37] reported juveniles of a species of Proteocephalus that possessed a vestigial muscular apical sucker, thus apparently not belonging to P. demshini , which is devoid of an apical sucker. T.Shimazu (unpubl. data) examined 100 specimens of B. toni from Hokkaido in 1984 and 2011 but no proteocephalidean cestodes were found, which indicates that P. demshini is endemic to the Primorsky Region of Russia.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |