
Plagiodontia aedium bondi, Turvey, Samuel T., Hansford, James, Kennerley, Rosalind J., Nuñez-Miño, José M., Brocca, Jorge L. & Young, Richard P., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3957.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:116FFA26-515E-49E8-9180-719B9B58F758 |
|
DOI |
https://doi.org/10.5281/zenodo.5682450 |
|
persistent identifier |
https://treatment.plazi.org/id/03A387AE-FFFC-FF9A-FF79-FBC8A63AE959 |
|
treatment provided by |
Plazi |
|
scientific name |
Plagiodontia aedium bondi |
| status |
subsp. nov. |
Plagiodontia aedium bondi subsp. nov. (James Bond’s hutia)
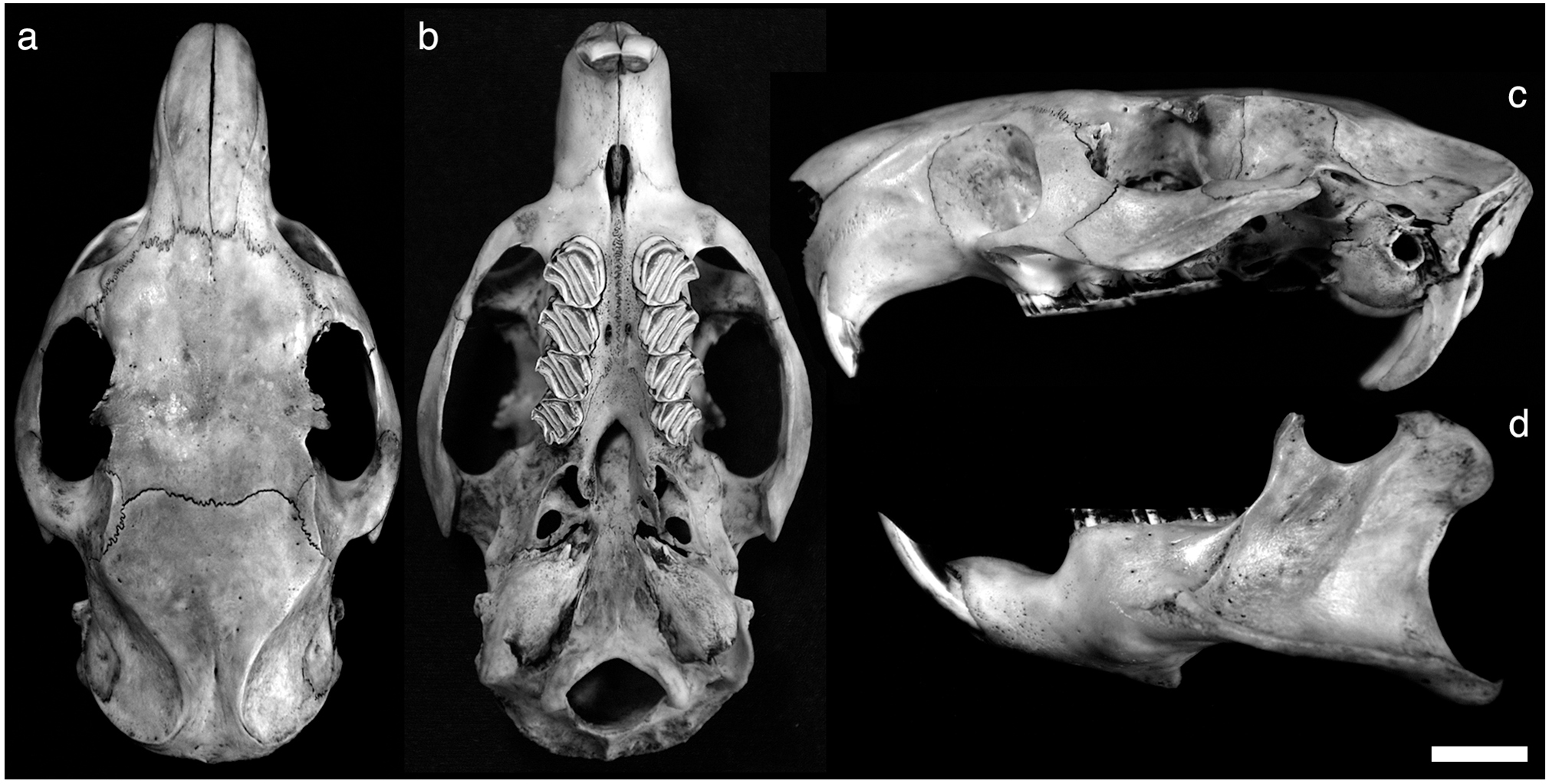
Holotype. LDUCZ-Z2807, complete skeleton of adult individual ( Figure 4 View FIGURE 4 ). Found dead in ravine, apparently after fatal fall from tree branch; collected by Ramon “Moncho” Espinal on 25 October 2011.
Paratype. LDUCZ-Z2808, complete skeleton of young adult with full tooth eruption and substantial but not complete closure of ectocranial sutures; possibly offspring of holotype individual. Collected at same location and date as holotype.
Etymology. Named after the ornithologist James Bond (1900–1989), to acknowledge his recognition of the biogeographic line in southern Hispaniola that represents the allopatric barrier between the new subspecies and P. aedium aedium .
Local names. This subspecies is referred to by local informants in southeastern Haiti and the southwestern Dominican Republic by several names, including jutía (commonest name used in Dominican Republic), kochondenn (commonest name used in Haiti), solenodon, kombee, and rata, reflecting confusion with other native and introduced co-occurring land mammals ( Turvey et al. 2014).
Type locality. Vicinity of Mencia village, Pedernales Province, Dominican Republic (18°10'10N, 71°44'25W), in area of primary broadleaf forest at elevation of 460 m.
Distribution. The southern palaeo-island of Hispaniola south of the Neiba Valley and east of the Jacmel– Fauché depression; found in both southeastern Haiti (departments Ouest and Sud-Est) and southwestern Dominican Republic (Barahona, Independencia and Pedernales provinces). Hutias are known to occur in the Massif de la Selle ( Haiti), and the Sierra de Bahoruco mountain range and Barahona Peninsula ( Dominican Republic) ( Sullivan 1983; Woods et al. 1985; Woods 1986; Woods & Ottenwalder 1992; Turvey et al. 2014).
Diagnosis. Subspecies of Hispaniolan hutia which is significantly larger than the closely related Plagiodontia aedium aedium in the following craniodental characters: maximum interorbital width, minimum anterior interorbital width, length and width of incisive foramina, external width across bony palate at PM4, crown width of PM4, M1 and M2, crown length of M3, crown width of pm4 and m1, coronoid–condyle length, and coronoid– angular length. Significantly larger than P. aedium hylaeum in width of incisive foramina and crown length of M2, and somewhat larger than this subspecies in crown length of M3, height of toothrow at pm4, and coronoid–condyle length. Postcranial and soft-tissue differences between all three subspecies are unknown.
Description. Craniodental anatomy of Plagiodontia aedium , including material from Pedernales Province referable to the new subspecies, described in detail by Woods & Howland (1979) and Woods (2001). Nasals slightly anteriorly inflated. No supraorbital ridge; slight postorbital process. Zygomatic arch broad, with greatest width opposite M3; well-defined lateral jugal fossa present, restricted to level of molar toothrow. Superior zygomatic root of maxillary broad, inferior zygomatic root narrow, with well-developed masseteric tuberosity on ventral surface; posterior zygomatic root formed by squamosal. Incisive foramen distinct, relatively short and broad; septum formed mainly by premaxillary. Alisphenoid canal forms crescent-shaped fissure in rear wall of orbit, with optic foramen small and deep within this fissure; no sphenopalatine foramen on medial canal wall. Posterior margin of palate with V-shaped mesopterygoid fossa extending opposite M3, and with long, delicate lateral pterygoid processes. Large pterygoid plate extends medially posterior to M3, contacting base of pterygoid process; masticatory foramen in middle of pterygoid plate; lateral pterygoid ridge absent. Pterygoid fossa restricted by pterygoid plate; medial to M3. Paroccipital process long and broad; closely associated with short, spike-like lateral process that is independent from bullae. Incisors not procumbent; upper incisors strongly curved; alveolar capsule of lower incisors terminates below m1. Molariform cheek teeth hypsodont and rootless; upper and lower toothrows both parallel. Cheek teeth with enamel ridges forming deeply overlapping cement-filled oblique reentrant folds oriented at 45° angle to body axis; upper cheek teeth with one labial re-entrant fold and one lingual reentrant fold; lower cheek teeth with one labial re-entrant fold and two lingual re-entrant folds. Anterior margin of upper cheek teeth sigmoid; lower cheek teeth not sigmoid. Mandible with well-developed coronoid process, and angular process with masseteric crest and broad, flat-bottomed pterygoid shelf; mental foramen absent; retromolar fossa present as large pit with mandibular foramen in its centre.
New subspecies distinguished phenotypically from other subspecies of P. aedium in this study on the basis of morphometric analysis using quantitative character measurements; no qualitative differences in craniodental character states are recognized. Craniodental measurements are as follows, taken from both individuals in type series unless indicated:
Skull: greatest occipitonasal length, 74.16–77.6 mm; height of skull above palate, 19.58–21.24 mm; height of braincase, 17.20–18.12 mm; width across braincase, 21.86–23.16 mm; width of occipital region, 27.76–28.22 mm; internal width across occipital condyle, 10.30–11.50 mm; width across paroccipital processes, 27.32–27.38 mm; height of occipital region excluding paroccipital processes, 17.56–17.60 mm; height of occipital region including paroccipital processes, 25.85–27.40 mm; width across auditory bullae, 28.74–29.05 mm; width across zygomatic arches, 41.00– 41.64 mm; minimum breadth of zygomatic plate, 4.38–4.88 mm; maximum interorbital width, 24.18–25.00 mm; minimum anterior interorbital width, 19.64–20.50 mm; minimum posterior interorbital width, 18.72–19.78 mm; maximum width of frontals, 23.58–25.50 mm; length of nasals, 25.10 mm (LDUCZ-Z2807 only); width of nasals, 9.20–9.60 mm; width of single incisor, 2.88–2.95 mm; length of diastema (from posterior incisor margin), 17.10–19.36 mm; length of incisive foramina, 7.00– 8.34 mm; width of incisive foramina, 2.80– 3.30 mm; internal width across bony palate at PM4, 2.54–2.60 mm; external width across bony palate at PM4, 14.74 mm (both specimens); internal width across bony palate at M3, 6.48–7.04 mm; external width across bony palate at M3, 15.55–15.96 mm; length from interior alveolus of PM4 to back of palate, 20.02–21.40 mm; width of mesopterygoid fossa, 4.24–4.56 mm; postpalatal length (to occipital foramen), 23.14–24.66 mm; crown length of cheek tooth row, 20.16–20.77 mm; alveolar length of cheek tooth row, 22.42–22.45 mm; maximum alveolar width of cheek tooth row, 5.94–6.58 mm; crown length of PM4, 6.18–6.26 mm; crown width of PM4, 5.36–5.85 mm; crown length of M1, 4.93–5.14 mm; crown width of M1, 5.75–5.96 mm; crown length of M2, 4.70–4.99 mm; crown width of M2, 5.30–5.62 mm; crown length of M3, 4.84–5.02 mm; crown width of M3, 4.58–4.64 mm.
Mandible: mandible length (incisor base to condyle), 50.64–50.98 mm; mandible length (incisor base to angular process), 51.58–55.34 mm; width of incisor, 2.70–2.78 mm; length of symphysis, 23.93–24.16 mm; length of diastema, 12.92–14.26 mm; length from incisor base to posterior bulb of incisor root, 21.52–22.34 mm; crown length of cheek tooth row, 19.09–20.16 mm; alveolar length of cheek tooth row, 21.92–21.96 mm; maximum alveolar width of cheek tooth row, 5.90–6.08 mm; crown length of pm4, 5.54–6.60 mm; crown width of pm4, 4.76–5.06 mm; crown length of m1, 4.76–5.00 mm; crown width of m1, 5.76–5.88 mm; crown length of m2, 4.32– 4.68 mm; crown width of m2, 5.46–5.62 mm; crown length of m3, 4.00– 4.36 mm; crown width of m3, 4.70–4.94 mm; maximum width of entire mandibular body, 21.00– 22.09 mm; maximum width of angular process, 6.02–6.95 mm; internal height of mandible (base to alveolar lip), 13.50–14.24 mm; internal height of mandible (base to tooth crown), 16.06–16.27 mm; height of toothrow at pm4 (internal aspect), 3.44–4.20 mm; height of toothrow at m3 (internal aspect), 1.12–1.20 mm; breadth across distal end of condyle, 10.32–10.37 mm; coronoid–condyle length, 15.74–18.36 mm; coronoid–angular length, 30.12–31.14 mm; condyle–angular length, 25.92–26.70 mm.
Detailed description of soft-tissue characters for the two specimens reported here is unfortunately not possible, as they were almost completely decomposed by the time they became available for study. General external morphology of hutias from southern Haiti, representing individuals of both P. aedium aedium and the new subspecies, was described by Woods & Ottenwalder (1992), and live individuals encountered in the Sierra de Bahoruco during fieldwork by the authors in 2009–2013 match the broad description given by these authors ( Figure 5 View FIGURE 5 a). However, previously published information on supposed differences in external body proportions and pelage characteristics between northern and southwestern Hispaniolan hutia populations ( Miller 1928; Johnson 1948; Anderson 1965; Woods & Howland 1979) is conflicting and of uncertain taxonomic value, and identification of systematic soft-tissue differences between different subspecies requires further study. Seventeen adult hutias were captured and released around Mencia in 2011–2013 during field research into ranging behavior and habitat use; adult males have a mean body mass of 1322.9 g (n=12, range= 1040–1795 g, SD=280.4), and adult females have a mean body mass of 1355.0 g (n=5, range= 1180–1530 g, SD=157.9), with total mean body mass for all individuals of 1335.0 g. These values are higher than the mean body mass of 1018 g for eight wild-caught individuals of P. aedium aedium from the Massif de la Hotte reported by Woods & Ottenwalder (1992).
Biological and ecological data. Hispaniolan hutias are considered to be predominantly arboreal when in goodquality forest ( Woods 1981; Sullivan 1983), but during the course of the 2011–2013 study some individuals were caught on the ground, and signs of feeding on saplings on the ground were often observed. Females caught on 6 January 2013 and 15 February 2013 were both pregnant, and a female caught on 14 September 2012 was lactating. Field observations and camera trap photos recorded young juveniles (200–350 g body mass) in separate family groups on 15 January 2012, 27 January 2012, 22 November 2012 and 11 January 2013, suggesting probable parturition during the autumn, with larger juveniles (420–500 g body mass) recorded on 12 May 2012 and 13 September 2012. Feeding signs detected during fieldwork and local reports suggest that hutias in the southeastern Dominican Republic feed upon a relatively wide variety of plants, including Rauvolfia (Apocynaceae) , Cleome (Cleomaceae) , Clusia (Clusiaceae) , Ipomoea (Convulvulaceae) , Tragia (Euphorbiaceae) , Acacia and Senna (Fabaceae) , Ocotea (Lauraceae) , Guarea and Trichilia (Meliaceae) , Ficus (Moraceae) , Trichostigma (Phytolaccaceae) , Gouania and Krugiodendron (Rhamnaceae) , Chrysophyllum and Sideroxylon (Sapotaceae) , and Guaiacum (Zygophyllaceae) .
The relatively limited existing literature on previous field studies of Hispaniolan hutias provides some further information on habitat preferences and ecological requirements of the new subspecies. Sullivan (1983) reported the presence of hutias referable to this subspecies in low-elevation (0–234 masl) dry subtropical forest around Laguna Oviedo on the Barahona Peninsula , where they have also been observed by the current authors ( Figure 5 View FIGURE 5 b), and also in humid subtropical forest in the Sierra de Bahoruco across an elevational gradient from mixed hardwood forest (500–900 masl) to deciduous vegetation in ravines within pine forest (1300–1500 masl), with all sites associated with rocky limestone substrate that provides suitable cavities and crevices for shelter. Hutias have also been recorded from the Massif de la Selle in southeastern Haiti in undisturbed broad-leaved forest (locally known as “rak bwa”) on the steep north slope of the main ridge (2160 masl) and along the ravine of the Riviere Blanche (1730 masl) ( Woods et al. 1985; Woods 1986; Woods & Ottenwalder 1992), again near areas of exposed limestone.
Interestingly, albinism has been documented in a relatively large number of individuals in this subspecies. Two separate albino individuals were caught during fieldwork around Mencia in 2011 and 2012, and a further albino individual was observed in Parque Nacional Jaragua in 2009 ( Figure 5 View FIGURE 5 b). A further “partially albinistic” individual “with pink eyes and unpigmented hands, feet and tail” from the “mountains of southern Haiti ” (specific location unknown), which could refer either to the Massif de la Hotte in southwestern Haiti or the Massif de la Selle in southeastern Haiti, was described by Tate (1948).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |