
Neacomys amoenus carceleni Hershkovitz, 1940
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4242.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:2DCFE9F6-7816-4CFC-ABAC-BC7A65AAF386 |
|
DOI |
https://doi.org/10.5281/zenodo.6022009 |
|
persistent identifier |
https://treatment.plazi.org/id/03C90206-9A7C-FFED-33BD-0B30FD48FED8 |
|
treatment provided by |
Plazi |
|
scientific name |
Neacomys amoenus carceleni Hershkovitz, 1940 |
| status |
|
Neacomys amoenus carceleni Hershkovitz, 1940
Alberto Carcelén’s Spiny Mouse
Neacomys spinosus carceleni Hershkovitz, 1940:1
Holotype. Adult male, skin, and skull U.M.M.Z. 80171, collected 5th October 1936 by Philip Hershkovitz, original number M618.
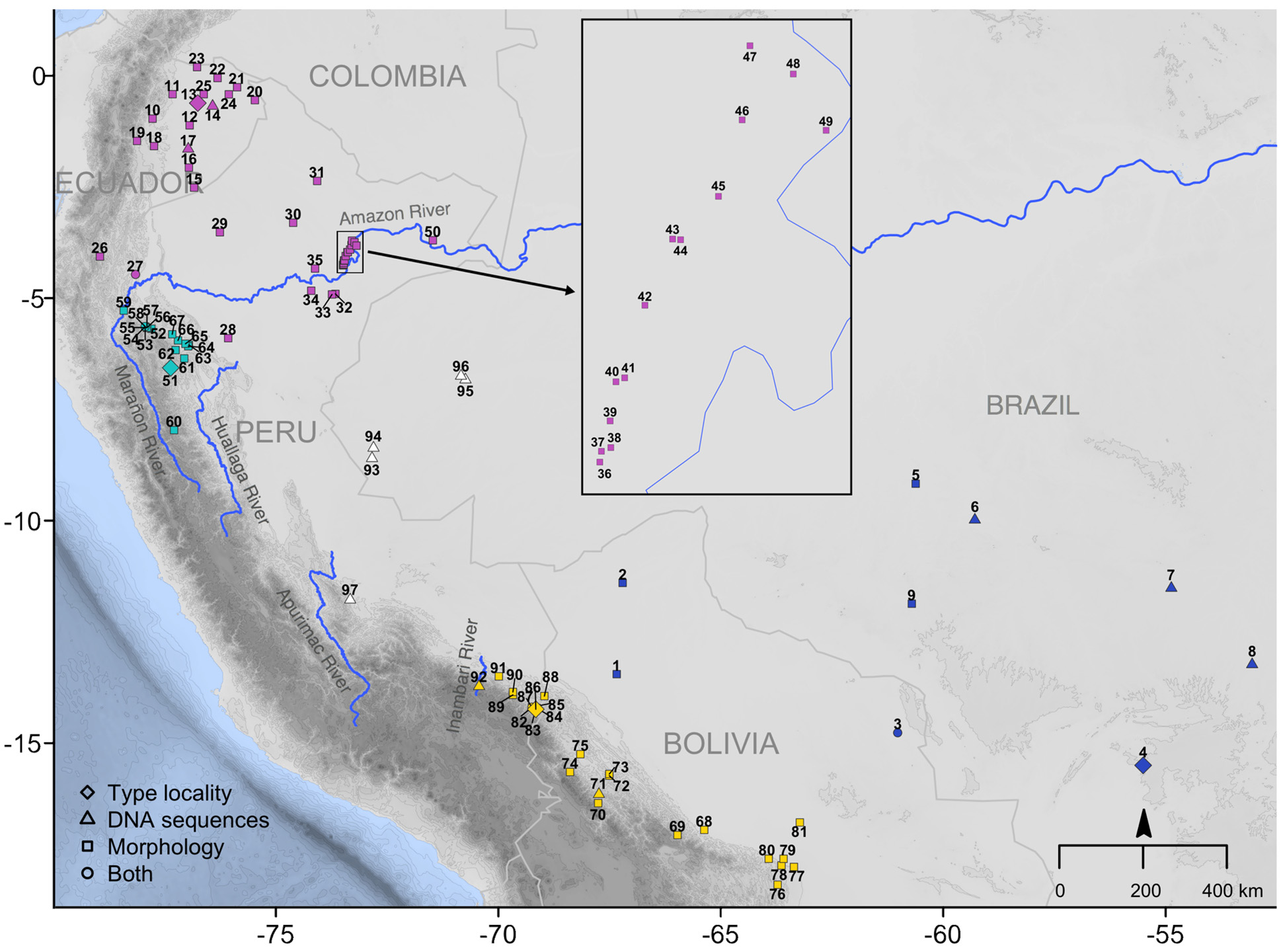
Type locality. “ Llunchi, an island on the northern side of the Rio Napo, west of the mouth of the Rio Jivino , latitude and longitude approximately 0°37’ S. , 76° 46’ W.; parish of La Coca, Napo-Pastaza Province , Ecuador; altitude about 250 meters.” Hershkovitz, 1940:1 ( Fig. 6 View FIGURE 6 ).
Emended diagnosis. Neacomys amoenus carceleni is different from other Neacomys because presents large body size, ventral fur completely white and short, long ungual manus tufts and carpal vibrissae, short hind feet, metatarsal and metacarpal patches always absent, tail subequal to head-body pencil of hairs is absent, rostrum broad and tapering with lateral humps, large gnathic process, squared lacrimals, well-developed interorbital beads, upper molar rows posteriorly convergent, diastema with a hump, anterior bullae process in contact with pterygoid plate, deep basioccipital pits, masseteric crest of the jaw below the hypoconid of the m1, deep crest between condylar and angular processes, anteroloph of M1 broad and asymmetrically divided, posteroloph of M1 large with a straight posterior border, and shallow protoflexus of M2.
Distribution. Distribution of Neacomys amoenus carceleni occurs in the Amazon basin, in Pastaza and Francisco de Orellana Provinces of Ecuador; Loreto, Madre de Dios and Ucayali Departments of Peru; and the Acre and Amazonas States of Brazil. Ranged from 200 to 750 m.
Morphological description. Large size. Dorsal fur is pale orange-brown; soft hairs are dark brown with pale orange tips, spines are white with gray tips; its length is 11 mm in average at the rump level; flanks are pale ochraceous. Ventral fur is completely white; it is short in length, 3 mm in average at the belly. Ungual manus tufts pass the claws. Fifth forefeet digit reaches the claw base of the fourth digit. Metacarpal patch is small, with colored hairs. Carpal vibrissae pass the second interphalangeal joint of the fifth digit. Genal and superciliary vibrissae pass the posterior pinna border. Basal protuberances for interramal vibrissae insertion are absent. Hind feet are slim and short, one-third larger than forefeet. Ungual pedal tufts do not pass the claws. First pedal digit passes the first interphalangeal joint of the second; fifth digit reaches the second interphalangeal joint of the fourth. Metatarsal patch is absent. Plantar squamation is on the pad and hypothenar region. Tail is subequal to head-body length. Upper tail hairs are black and cover 1–2.5 scales; lower hairs are whitish brown and cover 2–3.5 scales. Terminal pencil of hairs is absent.
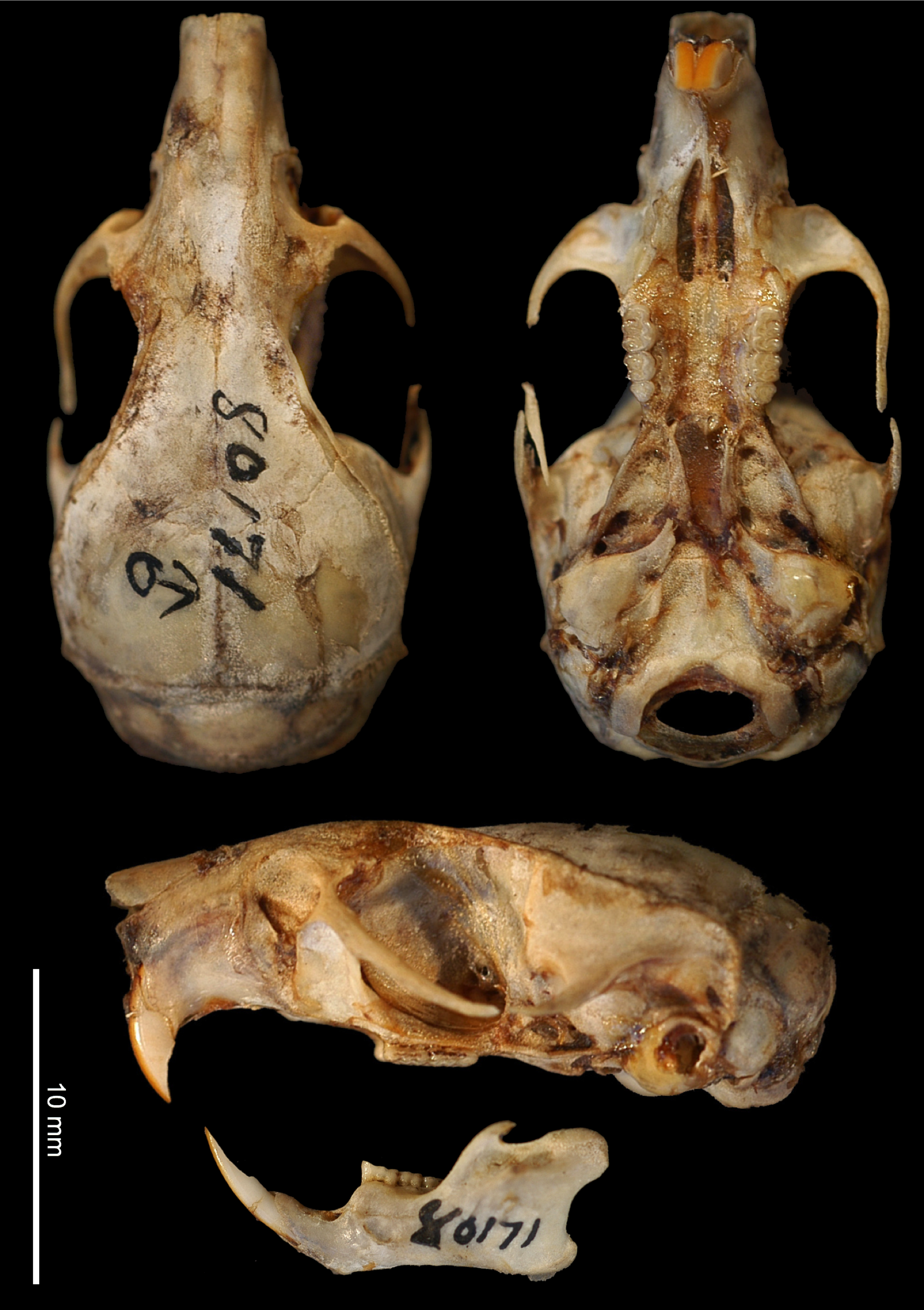
Skull. ( Fig. 9 View FIGURE 9 ) Dorsal view. Rostrum is broad and tapering, with lateral humps at premaxillary-maxillary suture level. Anterior borders of the nasals are rounded; sometimes exhibit a cleft between bones. Posterior borders of the nasals are irregularly rounded, behind premaxillary-maxillary-frontal joint and maxillary-frontal-lacrimal joints. Sutures between premaxillary-frontal are large and numerous. Zygomatic notch is shallow; does not enclose lacrimal foramen. Lacrimals are squared; one-half dip on squamosal root of zygomatic bone, and one-half in the frontal bone. Interorbital beads are well developed, but they are less developed in young individuals.
Lateral view. Skull profile is flat, but it exhibits a hump at frontal level. The most anterior part of the suture between nasal and premaxillary bones reaches the squamosal zygomatic root. Gnathic process is large, as the width of one upper incisive. Maxillary root width of zygomatic is as the M2 length. Zygomatic plate breadth is subequal to upper molar row length. Squamosal root breadth of zygomatic is one-half of zygomatic plate breadth. Ethmoid foramen is placed at the metacone of M1. Sphenopalatine foramen is placed at paracone of M2 level. Ethmoturbinals are rounded; they are as one-half of M3 length. Alisphenoid strut is always absent. Tegmen tympani are in contact with the posterior process of the squamosal. Malleus bone exhibits a large manubrium and a globose and large orbicular apophysis; the lamina of the malleus is rectangle and shallow. Processus brevis of incus is large, subequal to incus breadth, and well developed. Mastoid tubercle is not anteriorly concave. Upper border of postglenoid foramen is rounded. Subsquamosal upper border is tapered. Hamular process is thick. Mastoid perforation is present or absent, when is present it is posteriorly placed, with variable size and shape.
Ventral view. Inner borders of the incisive foramina are posteriorly parallel. Incisive foramina breadth is about M1-M2 length. Septum of incisive foramina is narrow. Diastema is not flat; presents a small hump. Origin of the superficial masseter is circular and flat. Maxillary-palatine suture is between hypocone and hypoflexus of M1 level. Posterior palatine foramina are at the protoflexus level of M2. Posterolateral palatal pits are placed from hypocone of M3 to anterior border of mesopterygoid fossa. Sphenopalatine vacuities are small or medium extended. Presphenoid is broad and with straight lateral borders. Mesopterygoid fossa breadth is lesser than incisive foramina breadth. Parapterygoid processes are convergent; laterally, they pass the molar surface level. Lateral borders of pterygoid plate are convex. Fontanellae are absent. Posterior opening of the alisphenoid canal is as M3 length. Groove for the infraorbital branch of the stapedial artery is shallow and wide. Middle lacerate foramina are narrow, as one-half of M2 length. Eustachian tube is large and thick. Bullas are globular. Anterior process of the bullae is in contact with the pterygoid plate. Basioccipital at petrosal level is broader than incisive foramina breadth. Basioccipital pits are deep. Anterior border of the foramen magnum is V obtuse shaped.
Jaw. Masseteric crest is below the hypoconid of the m1. Mental foramen is at the labial border of the diastema. Capsular process is moderately developed but does not pass the condylar process. Coronoid process is thick. Condylar process is short and broad. Angular process is short. Crest between condylar and angular processes are deep. Ventral border of the jaw is concave.
Teeth. Upper. Incisors are opisthodont, with rounded tips and golden-orange enamel bands. Molar rows are posteriorly convergent. Procingulum of M1 is anteriorly rounded, broad and asymmetrically divided. Anteroflexus of M1 is enclosed by the anterolabial and the anteroloph. Mesoloph is perpendicular to the labial surface. Posteroloph of M1 reaches the labial surface. Anteroloph of M2 is broad and do not reach the labial surface. Protoflexus of M2 is shallow. Mesoloph of M2 is perpendicular. Posteroloph of M2 does not reach the molar surface. Posterior border of the posteroloph of M2 is rounded. Lower. Procingulum of m1 is symmetrically divided. Mesolophid of m1 is narrow and do not reach the lingual surface. Hypoflexid of m1 is perpendicular to the labial surface. Posterior border of posterolophid of m1 is straight and reaches the lingual surface. Anterolabial cingulum of m2 is broad. Posterior border of posterolophid of m2 is straight and reaches the lingual surface.
Natural history. Neacomys amoenus carceleni occurs in the Amazonia from Peru, Ecuador and western Brazil going through Amazon and Marañon Rivers; inhabiting both secondary growth forests and cultivated areas ( Hice & Velazco 2012). Regarding reproduction, captured females were pregnant during dry season (between June and September) as in rainy season (between February and March), and lactating between November and December ( Hice & Velazco 2012); whereas males were recorded with scrotal testes in June, August, September, November, and December ( Hice & Velazco 2012).
Remarks. Hice & Velazco (2012) named individuals of N. a. carceleni as N. spinosus and provided a short description and figures of the skull, which agree with the diagnosis of N. a. carceleni ( Hershkovitz 1940; this study). Furthermore, Hice & Velazco (2012) recorded a candidate species from specimens caught with N. a. carceleni into the Reserva Nacional Allpahuayo-Mishana, and suggested that these species have a partition of habitats. The candidate species proposed by Hice & Velazco (2012) might be the specimens labeled as Neacomys sp. “clade 3” by Patton et al. (2000), due that two different clades were pointed out as co-distributed by these authors. Given that the names available in the synonymy of the genus are junior synonyms of N. amoenus s.l. (e.g. amoenus , carceleni ) and of N spinosus (e.g. typicus) (this study), the candidate species proposed by Hice & Velazco (2012) is still pending of description.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.