
Lissodendoryx (Ectyodoryx) corrugata, Fernandez, Julio C. C., Cárdenas, César A., Bravo, Alejandro, Lôbo-Hajdu, Gisele, Willenz, Philippe & Hajdu, Eduardo, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4092.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:51F44763-E817-4E58-A4AC-525E63B6D27B |
|
DOI |
https://doi.org/10.5281/zenodo.5615604 |
|
persistent identifier |
https://treatment.plazi.org/id/03EF87BE-FFCA-FFB7-419B-FF2AFEB7723D |
|
treatment provided by |
Plazi |
|
scientific name |
Lissodendoryx (Ectyodoryx) corrugata |
| status |
sp. nov. |
Lissodendoryx (Ectyodoryx) corrugata View in CoL sp. nov.
( Tabs 2–3 View TABLE 2 ; Figs 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Holotype. IZUA–POR 167, Isla Leucayec, Guaitecas Archipelago (44º03’59.00”S / 73º41’00.38”W, Chile), 10–18 m depth, coll. E. Hajdu & R. Foley, 0 7 March 2005. Fragments from holotype deposited under MNRJ 8963 and RBINSc–IG 32232–POR 8963. Paratype. MNRJ 17398, Punta Llonco, Comau Fjord, Chile (42º20’38.22”S / 72º27’25.26”W), <30 m depth, coll. G. Försterra, 0 3 January 2006.
Diagnosis. Massive, ovoid Lissodendoryx (Ectyodoryx) with numerous sinuous short anastomosing projections over the entire surface resembling a cauliflower; apically microspined tylotes (108–204/4.8–6), acanthostyles (I. 252–358/8–16.8, II. 90 –158/7.5–12.5), and arcuate isochelae (I 28–40, II 16–24).
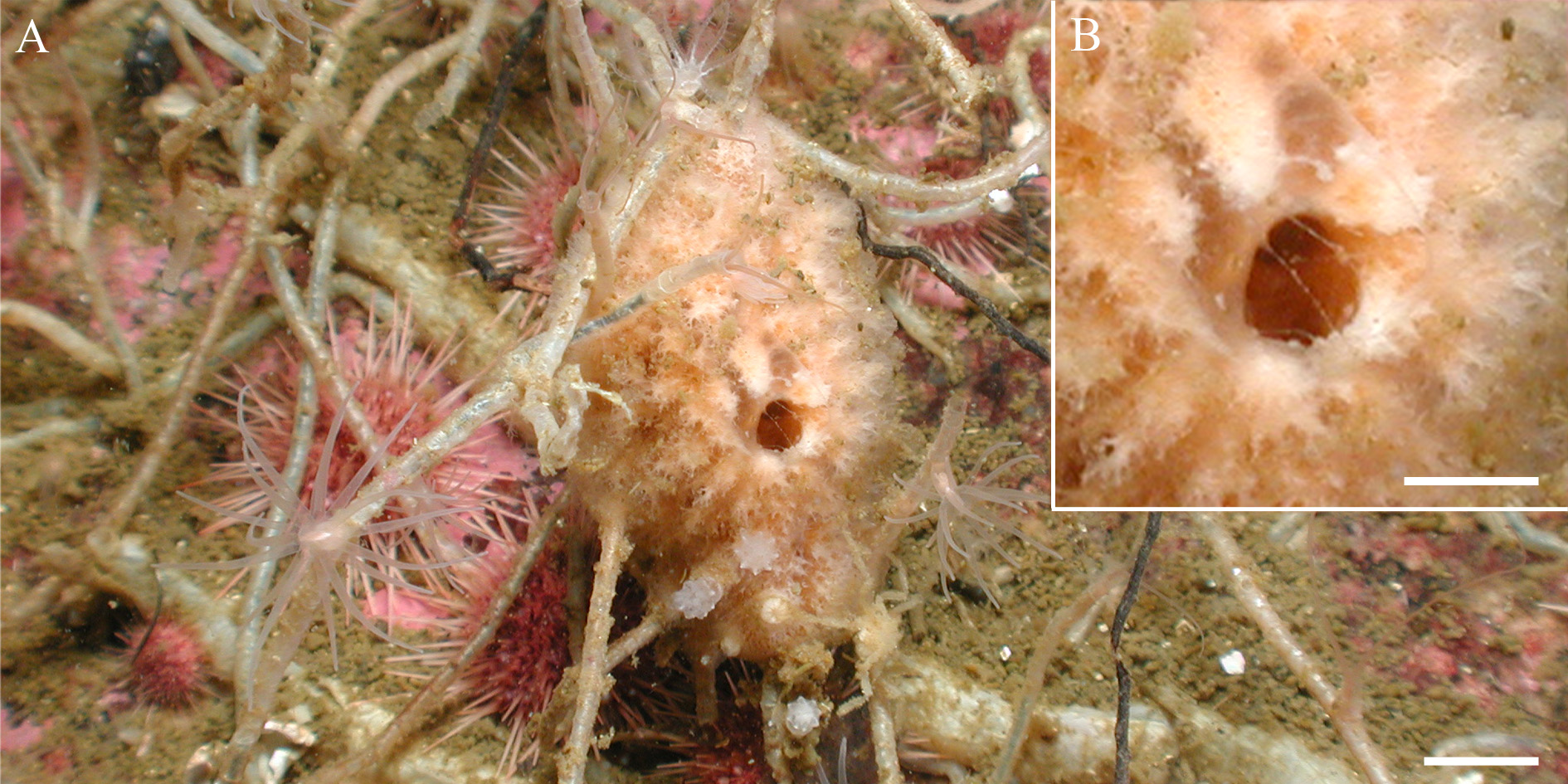
Description. Massive oval shaped sponge ( Figs 4 View FIGURE 4 A–B; Figs 5 View FIGURE 5 A–D), with numerous sinuous, short, anastomosing projections over the entire surface; resembling a cauliflower. The holotype is 4 cm long and 3 cm in high (in life) and the paratype is 3.8 cm and 3 cm, respectively. The paratype is relatively more compact. Simple oscula (diameter up to 0.3 cm, in vivo holotype), scattered and scarce. Colour in vivo beige, and in ethanol specimens are light beige. Their consistency is compressible, rather delicate, and the paratype is somewhat harder; texture slightly rough.
Skeleton. The choanosomal skeleton is (sub)anisotropic ( Fig. 6 View FIGURE 6 A) or subisodictyal reticulation ( Fig. 6 View FIGURE 6 D). Larger acanthostyles form pauci- to multispicular ascending tracts (up to seven spicules across), reaching the sponge surface and piercing it by up to 300 µm. These acanthostyles also constitute secondary orthogonal tracts, one spicule long, and up to four in thickness ( Figs 6 View FIGURE 6 B, E). Smaller acanthostyles echinate the main choanosomal tracts, and the nodes of the reticulation. Tylotes are spread in the surface, often perpendicularly or obliquely ( Fig. 6 View FIGURE 6 C). Tracts are partially inserted in a spongin layer of fibrous appearance ( Fig. 6 View FIGURE 6 F). Two categories of arcuate isochelae are scattered all around in choanosome and ectosome, the smaller of which is more frequent. Subectosomal lacunae absent, but wide choanosomal cavities occur, roundish or variably ellipsoid, up to 2 mm in maximum diameter.
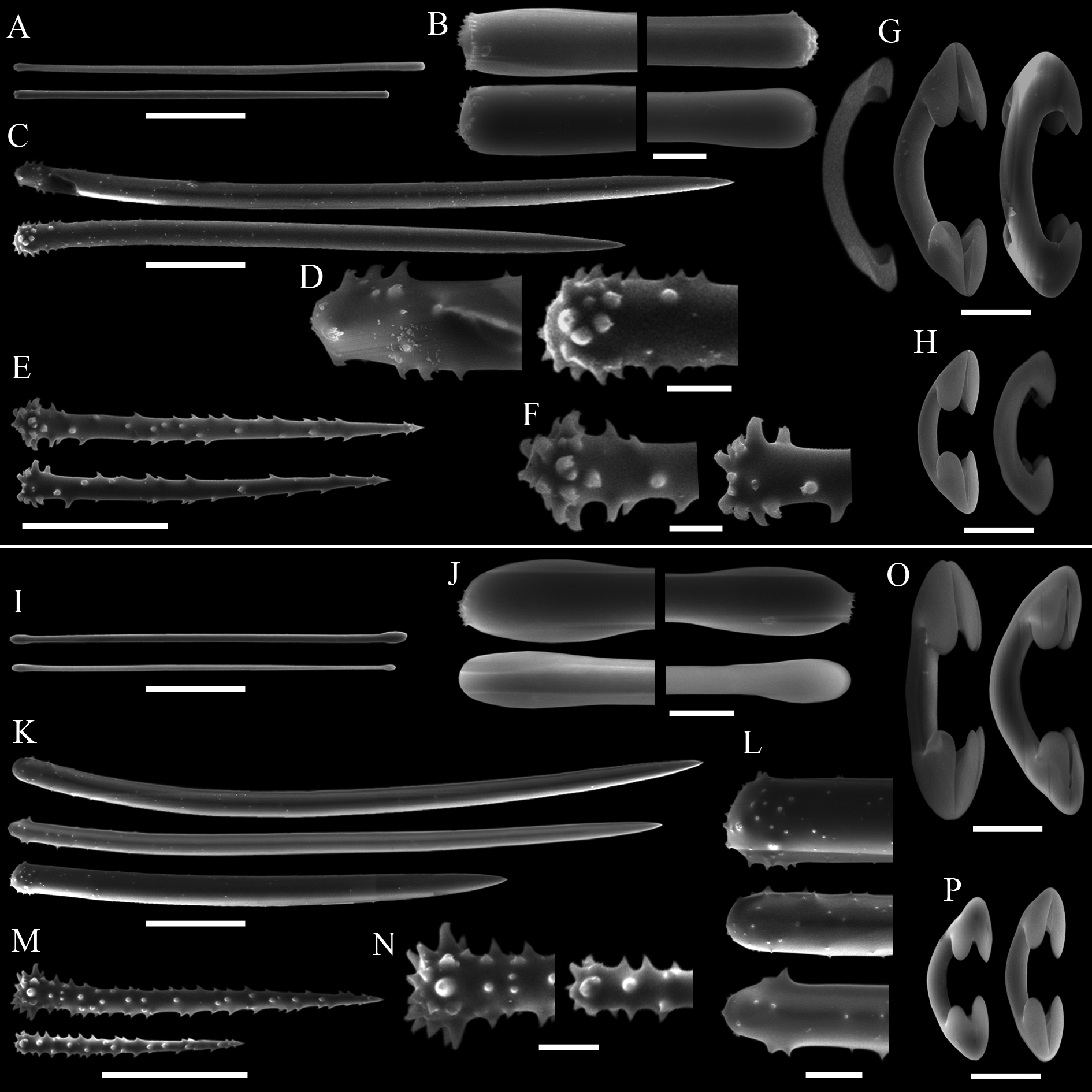
Spicules. Megascleres ( Tabs 2–3 View TABLE 2 ): (Sub)tylotes ( Figs 7 View FIGURE 7 A–B, I–J), straight, rather minutely microspined on both ends , which can be slightly aniso-tylote, elongated tyles only slightly swollen (elliptical), 108– 172 (25.3) – 204/4.8– 5.1 (0.3) –6. Acanthostyles I ( Figs 7 View FIGURE 7 C–D, K–L), straight or slightly curved, stout, somewhat fusiform, base slightly constricted, regularly round, apex sharpening gradually; spines not so abundant, straight, up to 1.5 µm high, concentrated at and near the base, a few spicules (variably thick) are very lightly spined or smooth, 252– 313.5 (29.7) –358/8– 13.5 (2.6) –16.8. Acanthostyles II ( Figs 7 View FIGURE 7 E–F, M–N), mostly straight, with a swollen base up to 3 µm thicker than the shaft, gradually sharpening point; abundant spines, up to 5 µm high, straight, spread over shaft and base, 90– 126 (24.3) –158/7.5– 10.3 (1.6) –12.5. Microscleres ( Tabs 2–3 View TABLE 2 ): Arcuate isochelae I ( Figs 7 View FIGURE 7 G, O), smooth, relatively thick shaft, alae slightly elongated, but relatively small, young forms with markedly reduced alae: 31– 34 (3.3) –40. Arcuate isochelae II ( Figs 7 View FIGURE 7 H, P), same as isochelae I, but smaller, 16– 22 (2) –29. The Sturges algorithm confirmed the occurrence of two size classes of isochelae.
Specimen ectosomal tylotes (with microspined choanosomal acanthostyles: arcuate isochelae
ends ) I. main, II. echinating
IZUA–POR 167 108– 162 –204/ I. 252– 300 –353/8– 14.7 –16.8 I. 31.5– 35.6 –40
holotype 4.8– 5.2 – 6 II. 90 – 128.2 –158/9– 11.2 –12.5 II. 16– 21 –24
MNRJ 17398 158– 182 –200/ I. 290– 326.6 –358/8.5– 12.3 –14.5 I. 28– 34 –40
paratype 4.8– 5 –5.2 II. 90 – 124 –145/7.5– 9.4 – 10 II. 21 – 22.8 –24 Ecology. The holotype was attached to a bunch of slender chitinous polychaete tubes (Family Spionidae ), and the paratype was attached to a coral.
Distribution. So far endemic from the northern sector of Chile’s fjord region, from its type locality at the Guaitecas Archipelago (44ºS) to Comau Fjord (42ºS).
Etymology. The species is named ‘corrugata’ (Latin corrugatus = rugose) on account of its irregular, cauliflower-like surface.
Remarks. Lissodendoryx (Ectyodoryx) corrugata sp. nov. is distinguished from Lissodendoryx (E.) spp. occurring in the SE Pacific, and in additional allied biogeographic provinces, as well as from L. (E.) ballena sp. nov. (described above) by its possession of two categories of arcuate isochelae combined with terminally microspined tylotes. The presence of little spines in the extremities of tylotes is shared with five species of Lissodendoryx (Ectyodoryx) considered here ( Tab. 2 View TABLE 2 ); viz. L. (E.) anacantha , L. (E.) nobilis , L. (E.) patagonica , L. (E.) plumosa , and L. (E.) ramilobosa . This character may also be present in the genus’ type species, L.
(Lissodendoryx) isodictyalis (Carter, 1882) . A phylogenetic assessment of synapomorphies is needed to verify whether the current subgeneric arrangement, and its emphasis on presence vs. absence of echinating acanthostyles (van Soest, 2002a), is more parsimonious than an alternative system with greater weighting given to the micromorphology of spicules.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Lissodendoryx |