
Limnonectes beloncioi, Herr & Goyes Vallejos & Meneses & Abraham & Otterholt & Siler & Rico & Brown, 2021
|
publication ID |
https://doi.org/ 10.1643/h2020095 |
|
DOI |
https://doi.org/10.5281/zenodo.7850057 |
|
persistent identifier |
https://treatment.plazi.org/id/0E16396A-5921-FFDF-2A98-6458F6A6FC0C |
|
treatment provided by |
Felipe |
|
scientific name |
Limnonectes beloncioi |
| status |
sp. nov. |
Limnonectes beloncioi , new species
urn:lsid:zoobank.org:act:4CFCEF76-99D6-40DC-9B53-58DECB1027CF
Mindoro Fanged Frog
Figures 1 View FIG , 6 View FIG , 7 View FIG
Rana macrodon blythi Boulenger, 1920 (partim). Rana acanthi Taylor, 1923 ; Taylor and Elbel, 1958. Rana macrodon acanthi ( Inger, 1954) . Rana macrodon macrocephala ( Inger, 1954) (partim: three
Mindoro specimens [USNM] provisionally referred by
Inger’s ‘‘tentative identification’’ [ Inger, 1954: 129]). Rana magna acanthi (Inger, 1958) .
Limnonectes (Limnonectes) acanthi Dubois, 1987 View in CoL (partim). Limnonectes cf. acanthi Evans et al., 2003 View in CoL ; Setiadi et al., 2010;
Diesmos et al., 2015.
Holotype.— PNM 9870 (adult male; formerly KU 303343 ; Field collector No. RMB 4957), Philippines, Mindoro Island, Oriental Mindoro Province, Municipality of Bongabong, Barangay Carmundo , Sitio Paypay-Ama , Paypay-Ama River , 12.7354 ° N, 121.4141 ° E, 100 m above sea level, WGS 84, R. M. Brown, A. C. Diesmos, C. D. Siler, and E. L. B. Rico, 13 March 2005. GoogleMaps
Paratypes (Paratopotypes).— KU 302084 , 302087–88 (adult females), 302085–86 , 302089 (adult males), 303343 (juvenile of undetermined sex), 303369–78 (10 subadults of undetermined sex), bearing the same data as the holotype.
Other paratypes.— Mindoro Island, Oriental Mindoro Province, Municipality of Bongabong, Barangay Formon : KU 302090–91 (adult females), 302093 , 302095 , 302097 , 302100 (3 adult males, 1 female), 302109–11 (3 adult males), C. D. Siler, 12 March 2005; Municipality of Victoria, Barangay Loyal : KU 302112–18 (2 adult males, 2 adult females, 3 juveniles of undetermined sex), C. D. Siler, 13 March 2005; Barangay Loyal, Sitio Panguisan, Panguisan River : KU 303470–78 (4 adult females, 5 subadults of undetermined sex), R. M. Brown, A. C. Diesmos, and C. D. Siler, 14 March 2005; Municipality of Gloria, Barangay Malamig : KU 302108 (adult female), 303344 (juvenile), J. B. Fernandez and R. M. Brown, 17 March 2006; KU 303346–54 (2 adult males, 2 females, 5 juveniles), R. M. Brown, C. D. Siler, and A. C. Diesmos, 13 March 2005; Sitio Balogbog, Cueba Simbahan : KU 303379–80 (2 subadults of undetermined sex), R. M. Brown, C. D. Siler, and E. L. B. Rico, 12 March 2005; Sitio Pastohan, Tanguisian Falls : KU 303381–402 (22 subadults of undetermined sex), A. C. Diesmos and E. L. B. Rico, 11 March 2005; Occidental Mindoro Province, Municipality of Calintaan, Barangay New Dagupan: KU 303266 , 303345 (subadults), R. M. Brown, 8 March 2005; Municipality of Magsaysay, Barangay Nicolas, Sitio Banban : KU 303404–30 (1 adult female, 25 subadults and juveniles /metamorphs of undetermined sex), C. D. Siler and R. M. Brown, 9 March 2005; KU 304131–32 (adult male and subadult of undetermined sex), R. M. Brown; Municipality of Sablayan, Barangay Batong Buhay, Sitio Batulai , Mt. Siburan : KU 303430–52 (5 adult males, 6 adult females, 12 subadults of undetermined sex), E. L. B. Rico, 14 February 2006; KU 305450–51 , 306637 (adult female, 2 subadult females), E. L. B. Rico, 19 February 2006; Barangay Malisbong, Sitio Aruyan : KU 335863–83 (11 females, 10 males), S. N. Travers, C. H. Oliveros, and R. M. Brown, 6 July 2013; Barangay Burgos, Sitio Posoy, Posoy River : KU 303453–69 (adult male, adult female, 15 juveniles of undetermined sex), R. M. Brown, 8 March 2005; Municipality of Paluan, Barangay Harrison, Sitio Ulasan , local name ‘‘ Matingaram’ ’: KU 308307 , 308309 , 308313–18 , 308321–23 , 308327 , 308360 , 308362–63 , 308367–68 , 308370–71 , 308385 , 308391 , 308393 , 308422 , 308457 , 308462 , 308464–65 , 308469 , 308472 (15 adult females, 15 adult males), E. L. B. Rico, 4 January 2007; Municipality of Puerto Galera, Barangay San Isidro, Sitio Minolo , Ponderosa Golf Resort , adult female, J. A. McGuire and V. Yngente, 15 January 1996: TNHC 54920 ; Municipality of San Teodoro, Barangay Villaflor, Tamaraw Falls , approximately km 15 from Puerto Galera on Calapan-to-Puerto Galera road, 8 subadult males, 1 immature female, 3 adult males, J. A. McGuire and V. Yngente, 17 January 1996: TNHC 54921–29 , 55023 , 55025 , 55029 , 55033 ; same locality, 10 adult males, 11 adult females, R. I. Crombie and V. Yngente, 8 March 1995: USNM 556073–94 ; Municipality of Baco, Barangay Lantuyan, near Cabinuangang River : 6 adult males, R. I. Crombie and V. Yngente, 2 July 1991: USNM 508558–63 ; 5 adult males, 3 adult females, 1 immature specimen of undetermined sex, R. I. Crombie and V. Yngente, 7 March 1995: USNM 508564–72 ; Municipality of Tarogin, ca. 30 km S of Calapan Town, Mt. Halcon SE slope: CAS-SU 22146 (adult female), Q. Alcala and party, 1 April 1963; CAS-SU 22145 (adult female), same data, 31 March 1962; CAS-SU 22147–49 (adult male, 2 adult females), 1 April 1963; CAS-SU 22150 (adult female), S. Magusara and C. Batal, 14 April 1963; CAS-SU 22576 (adult male), Q. Alcala and party, 13 March 1963; CAS-SU 22577 , 23508 (adult male and female), 31 March 1963; CAS-SU 23499 , 23501 , 23525 (adult females), 23505 , 23514–15 , 23519–20 (adult males), CAS-SU 23485 , 23487 , 23496–97 , 23512–13 , 23522 (subadult males), 23489 , 23498 , 23502 (subadult females), Q. Alcala and party, 10 March 1963; Municipality of Tarogin, Mt. Halcon : CAS-SU 22240 (juvenile), Q. Alcala and party, 14 April 1963; CAS-SU 22288– 22295 , 23500 , 23510–11 , 23517–18 , 23521 (juveniles), Q. Alcala and party, 1–20 April 1963; E side of Mt. Halcon, SE slope of Barawanan Peak, 830 m: CAS-SU 22151 (adult female), M. Pinero and party; Semirara Island, Oriental Mindoro Province, Municipality of Caluya , Barangay Tinogboc : KU 302105–07 (2 adult males, 1 adult female), C. D. Siler, 16 November 2000.
Referred specimens.— Mindoro Island, Oriental Mindoro Province, Municipality of Baco, Mt. Baco , Alangsa River : USNM 508534–57 ; Occidental Mindoro Province, Municipality of Paluan , Barangay Harrison, Sitio Ulasan , local name ‘‘ Matingaram’ ’: KU 308308 , 308310–12 , 308319–20 , 308324–26 , 308361 , 308364–66 , 308369 , 308372–76 , 308386–90 , 308392 , 308394 , 308416–21 , 308423 , 308430 , 308451–52 , 308456 , 308461 , 308463 , 308467–68 , 308470–87 , 308500 , 308528 , 308538 , 308561–69 , 308586 , 308589 , 308590–92 ; Municipality of Paluan, Barangay 1 , Sitio Ipol : KU 308593 , 308597 , 308599 .
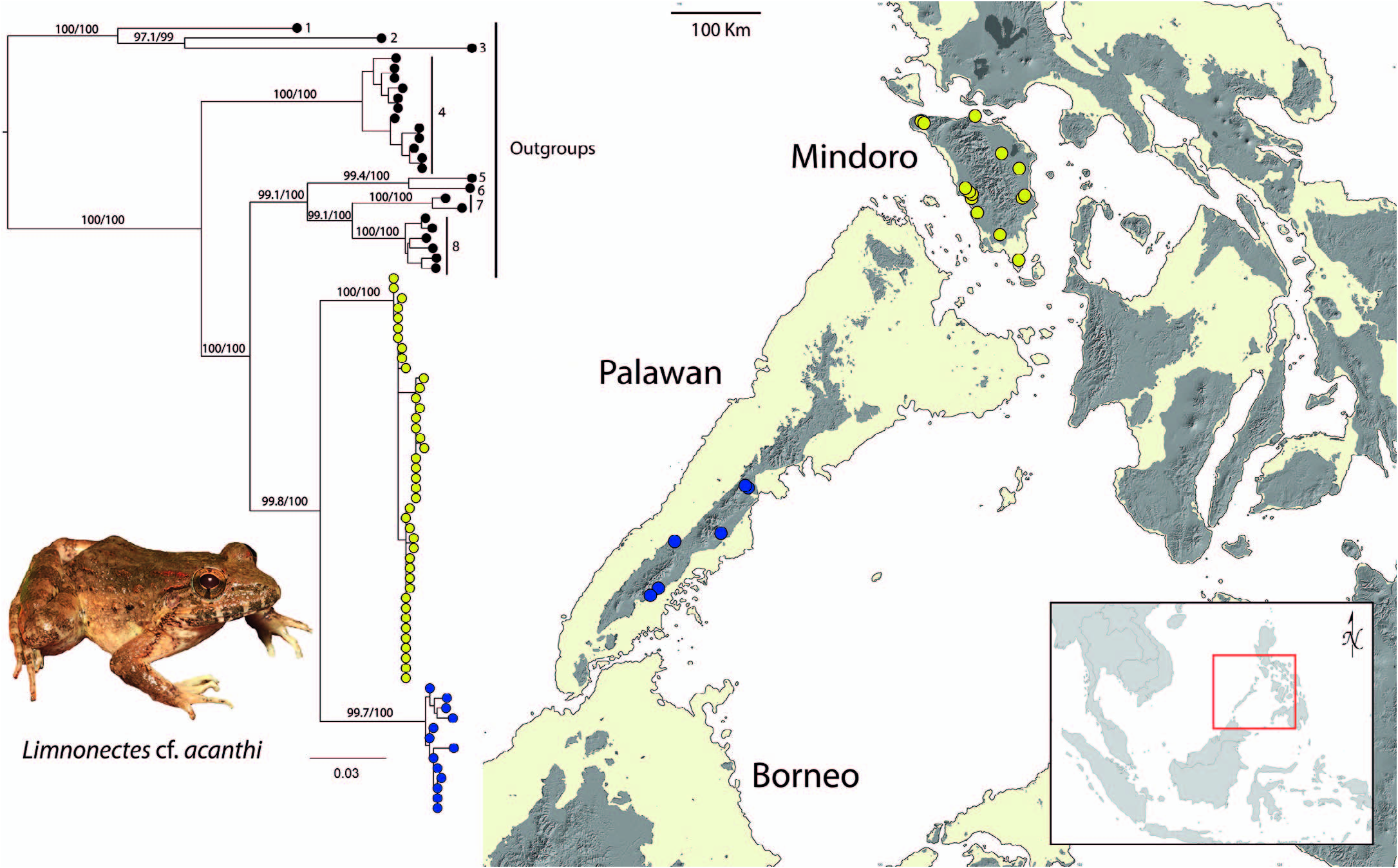
Diagnosis and comparisons.— Limnonectes beloncioi is a medium-sized fanged frog, assigned to the genus Limnonectes (family Dicroglossidae ), on the basis of its prominent, sexually dimorphic odontoid processes characteristic of the genus among other osteological synapomorphies ( Inger, 1954, 1966; Emerson and Berrigan, 1993). The new species can be distinguished from all other known congeners based on a combination of its single-pulse/note advertisement call (vs. dual-pulses/note in L. acanthi from Palawan Island faunal region and PAIC landmasses), its phylogenetic position (sister to L. acanthi from Palawan PAIC; Evans et al., 2003; Setiadi et al., 2011), and its geographic distribution on Mindoro and Semirara Islands (vs. Palawan PAIC); it is the only species of Limnonectes known to occur on Mindoro and Semirara Islands and associated small satellite islands and, therefore, has no sympatric congeners.
The new species is morphologically similar to its closest relative, L. acanthi ; however, it may be distinguished from this allopatric congener by its male advertisement call. The note pulse substructure of L. beloncioi is singular (1 pulse per note vs. 2 pulses per note in L. acanthi ), and the new species has a slower note repetition rate (11.5–12.8 notes per second vs. 14.2–15.4 notes per second), and has a higher dominant frequency (2,067.2–2,799.3 Hz vs. 1,335.1–1,679.6 Hz in L. acanthi ).
With the exception of the morphologically indistinguishable L. acanthi , the new species can be distinguished from all other Philippine species of Limnonectes ( L. diuatus , L. ferneri , L. leytensis , L. macrocephalus , L. magnus , L. micrixalus , L. palavanensis , L. parvus , L. visayanus , and L. woodworthi ) by a combination of body size, fang (odontoid) length, snout shape, relative lengths of the first and second finger, dorsal skin rugosity, restriction of white-tipped dermal asperities to the sacral region (not aggregated in radial clusters), the presence of irregular, elongate, discontinuous dorsolateral ridges (absence of a continuous dorsolateral fold), complete interdigital webbing of the foot, and the absence of a dark inverted ‘‘V’’-shaped mark on the dorsum. We provide morphological comparisons below, based on our data, in conjunction with or with consideration of the descriptions of Stejneger (1910), Taylor (1923), Inger (1954), Brown and Alcala (1977), and Siler et al. (2009).
Limnonectes beloncioi differs from L. diuatus and L. ferneri by its rounded snout in lateral aspect (vs. posteroventrally sloping), its Finger I> Finger II relative finger lengths (vs. approximately equivalent length), and by restriction of white-tipped dorsal asperities to dorsal sacral region, and not distributed in radial clusters (vs. asperities not posteriorly restricted in L. diuatus ; and densely distributed across entire of dorsum, and concentrated in radial sacral clusters in L. ferneri ); and the presence of irregular dorsolateral ridges (absent in L. diuatus and L. ferneri ); from L. leytensis , the new species can be distinguished by its larger adult body size (SVL range 54.2–83.1 vs. 25.8.2–34.0 mm in L. leytensis ), rounded snout (vs. snout moderately pointed in lateral aspect in L. leytensis ), Finger I> Finger II (equivalent length in L. leytensis ), complete webbing (webbing incomplete/reduced in L. leytensis ), and the absence of an inverted ‘‘V’’-shaped mark on the anterior dorsum (vs. present in L. leytensis ); from a large male specimen of L. macrocephalus (the species endemic to the Luzon PAIC landmasses of Luzon, Polillo, Catanduanes, and Marinduque), the new species can be distinguished by the observation that it attains a considerably smaller maximal adult male body size (SVL range 54.2– 83.1 vs. 78.9–144.6 mm in L. macrocephalus ); the new species also possesses a fully exposed tympanum (vs. dorsal and/or posterior edge of tympanum hidden beneath overlapping supratympanic dermal ridge skin in L. macrocephalus ) and lacks sexual size dimorphism ( Table 2 View Table 2 ), whereas L. macrocephalus exhibits reverse sexual size dimorphism (males larger); from a large adult male specimen of L. magnus , the new species can similarly be distinguished by its smaller adult body size (SVL range 54.2–83.1 vs. 66.3–164.4 mm in L. magnus ), rugose middorsal skin texture (vs. smooth to shagreened in L. magnus ), fully exposed tympanum (vs. dorsal and/or posterior edge of tympanum hidden beneath overlapping supratympanic dermal ridge skin in L. magnus ), rounded snout (vs. pointed in L. magnus ), and by the absence of reverse sexual size dimorphism (present in L. magnus ); from L. parvus , L. micrixalus , and L. palavanensis , the new species can be distinguished by its larger adult body size (SVL 54.2–83.1 mm; vs. 24.2–35.5 in L. parvus ; 28.1–30.2 in L. micrixalus ; 30.0–37.6 in L. palavanensis ), rugose (vs. smooth) dorsal skin, Finger I> Finger II (equivalent length), the presence (vs. absence) of white-tipped sacral asperities and the presence of irregular, discontinuous dorsolateral ridges (vs. asperities absent, dorsolateral folds continuous), complete webbing (vs. reduced), and by the absence (vs. presence) of an inverted ‘‘V’’-shaped middorsal marking (vs. present); the new species is additionally distinguished from these species by the absence of sexual size dimorphism (vs. females larger in L. parvus , L. micrixalus , and L. palavanensis ); from L. visayanus , L. beloncioi is readily diagnosed by its rounded snout (vs. pointed in L. visayanus ), a tendency towards longer adult male fangs (2.6–5.6 vs. 1.8–3.0 mm in L. visayanus ), by the presence (vs. absence) of white-tipped sacral asperities; finally, the new species can be distinguished from L. woodworthi by its longer male fangs (2.6–5.6 vs. 1.3– 1.6 mm in L. woodworthi ), moderately rugose middorsal skin (vs. smooth in L. woodworthi ), rounded snout (vs. snout moderately pointed in L. woodworthi ), the presence (vs. absence) of white-tipped sacral asperities, and by the absence (vs. presence) of continuous dorsolateral folds.
Description of holotype.— A mature male, specimen in excellent condition; small portion of liver preserved separately for genetic material; habitus robust; head broader than body, its length 98.3% of its width, 39.0% of SVL; snout tip rounded in dorsal and lateral aspect ( Fig. 6 View FIG ); supralabial region markedly swollen, increasingly protuberant towards angle of jaw; interorbital region and dorsal rostrum nearly flat; eye diameter 62.0% snout length, 97.5% eye–nares distance, 1.4 ⅹ eye–tympanum distance; pupil horizontally sub-elliptical with discontinuous posterior margin; canthus rostralis distinct, slightly medially bowed in dorsal aspect; loreal region concave; nostrils oriented dorsolaterally, narial openings visible in dorsal view; internarial region slightly convex; tympanum exposed, annulus slightly distinct, diameter 55.7% of eye diameter; supratympanic fold thick, strongly protuberant, moderately rugose, extending from posterior corner of eye, extending horizontally over (concealing) dorsoposterior corner and posterior margin of tympanic annulus, turning ventrally at nearly right angle, to end in supra-axillary region, where it is discontinuous with postrictal tubercular swelling at angle of jaw. Tongue elongate, tapered anteriorly, with narrow anterior attachment and laterally expanded, free, bilobed posterior margin at rest (anterior edge when tongue projected); choanae situated at anterolateral edge of palate, subcircular, their anterolateral edge partially concealed by palatal shelf of maxilla in ventral view; choanae widely separated by distance five or six times greater than diameter of single choana, each located just anterolaterally to (in contact with) lateral tip of dentigerous process of vomer; dentigerous process of vomer distinct, with four or five conical teeth on each side; dentigerous process angled anterolaterally (rostrally), approximately at 45 ° inclination, with closest (posterior) points separated by distance approximately equal to one choana, their most distant (anterior) ends separated by distance equal to three choanae; enlarged odontoid ‘‘fangs’’ large, recurved, unsheathed by oral mucosa for> distal half their length, situated on either side of mandibular symphysis/medial bulge, their tips sharply pointed, total length 4.6 mm (perpendicular distance from ventral edge of mandible), inclined dorsoposteriorly, tips 2.9 mm perpendicular from dorsal mandible surface; maxillary fang ‘‘sockets’’ anteromedial to choanae, large, round, similar in size to one choana; vocal apertures large, elongate, surrounded by extensive mucosal invaginations, situated at posteroventral margin of buccal floor, just medial to angle of jaw.
Hand length 46.8% foot length; foot 93.4% tibia length; tibia length 58.3% SVL; fingers laterally, irregularly ovoid in cross section, due to presence of slight lateral dermal flange, extending from base of each digit, on either side, to proximal margins of terminal finger discs; terminal discs not expanded beyond widths of penultimate phalanges ( Fig. 6 View FIG ), their relative descending lengths: III> I> II ¼ IV; subarticular tubercles prominent, their ventral surfaces convex and velvety in texture; one subarticular tubercle below Fingers I and II, two tubercles under Fingers III and IV; terminal discs and subarticular tubercles with gray, velvety, thickened surfaces; distal margins of tubercle more distinct than their proximal margins, and supernumerary tubercles absent, but articular surfaces of fingers between subarticular tubercles of digits, and at base of all digits covered medially with thickened tubercular surface; palmar surface with large, elongate, thenar tubercle (ventromedial surface of Finger I), enlarged, flattened, squarish medial ‘‘inner’’ (base of Finger III) metacarpal tubercle, and small, ovoid, convex outer (base of Finger IV) metacarpal tubercle; surface of these palmar structures, intervening, and surrounding surfaces all covered with similar, thickened, velvety (matte) tubercular dermis layer; nuptial excrescences or pads, asperities, and webbing absent; forearm musculature not hypertrophied.
Tarsus folds and flaps absent; terminal discs of toes slightly expanded, with distinct circummarginal grooves; plantar surfaces of foot with well-developed, prominently protruding (ventrally), rounded subarticular tubercles ( Fig. 6 View FIG ); plantar surfaces of foot smooth, with velvety-textured subarticular tubercles; relative lengths of toes: I <II <V <III <IV; toes fully webbed (interdigital webbing extending to proximal edges of terminal discs of all toes); postaxial flap of skin running along entire outer edge of Toe V; inner metatarsal tubercle prominent, elongate, with raised ventral edge; outer metatarsal tubercle absent.
Skin of dorsal surfaces of trunk and head smooth to slight shagreened texture, bearing low but clearly evident fleshy dermal tubercle clusters in supra- and post-tympanic regions, and tuberculate dorsolateral ridges, immediately following supratympanic region, and continuing posteriorly to the scapular region approximately to the points of forearm insertion ( Fig. 6 View FIG ); similar dorsolateral tubercular ridges extend from this point, along more lateral (flanks) and slightly medial (dorsal) surfaces, and extend back to the sacral region; on posterior half of trunk and sacral region, dermal tubercles present mid-dorsally, consisting of single, raised tubercles or short, raised tubercular ridges; in sacral region, some tubercles capped with round, weakly keratinized dermal asperities; not arranged in clusters, or rows, but lightly dispersed in sacral region and upper one-third of dorsal surface of thigh and supra-cloacal region; ventral surfaces of head smooth; lateral and ventral surfaces of limbs smooth; remaining dorsal surfaces of limbs smooth to lightly shagreened, with occasional low tubercles; tarsus smooth on dorsolateral surface; cloacal region rugose (wrinkled), with smooth laterally and ventrally surrounding skin.
Coloration of holotype in preservative.— Dominant dorsal color on head, body, and forelimbs uniform Dark Grayish-Brown ( Köhler, 2012; color 284) Dark Grayish-Olive (275) with irregular, diffuse, Dusky Brown markings (285) concentrated on occiput and sacral regions; lateral head surfaces Drab (19) with diffuse Warm Sepia (40) markings, canthal bar, pigmentation associated with supratympanic ridge; four Medium Neutral Gray (298) labial bars alternate with lighter, Pale Buff (2) labial region coloration; Sepia bar (286) spans the interorbital region; tympanum Dark Gray (299) with Pale Neutral Gray (296) central spot; dorsal surfaces of hindlimbs Brussels Brown (33) to Brick Red (36), with darker, Sepia (286) diffuse transverse crossbars; dorsal surface of outer (Finger IV) hand and (Toes III–V) Drab Gray (256); inner dorsal surfaces of hand (Fingers I–III) and Toes (I–II) Bright White Buff (a) to Cream White (52); ventral body and proximal limb segment surfaces Cream White (52) to Chamois (84), nearly Cream Yellow (82) on upper ventral thigh surfaces and infracloacal region; sternal region and throat with dense aggregation of distinct Orange-Rufous (56) blotches; infralabial region Grayish Horn (268), mandibular region Cream White (52); ventral surfaces of hand and foot Vandyke Brown (181) to Glaucous (291) with Jet Black (300) ventral palmar surfaces, post-brachial surfaces of forearms, and ventral surfaces of tarsal (shank) surfaces; central surfaces of subarticular tubercles of hand and foot Light Neutral Gray (297) and terminal discs of fingers and toes Pale Neutral Gray (296).
Coloration of holotype in life.— Based on field notes and color images by RMB. Dorsal surfaces light brown (Sayal to Mikado browns, colors 41–42) with Burnt Umber Brown (48) and Grayish Horn (268) diffuse, irregular, scattered darker markings and Jet Black pigmentation associated with dorsolateral and lateral flank tubercles, the supratympanic ridge, postrictal coloration, and labial bars; dorsal surfaces of limbs Sayal Brown (41), Prout’s Brown (41), to Natal (49) dark brown blotches and thick transverse limb bars; lateral head with Dark Neutral Gray (299) canthal, snout, and post-ocular pigment; tympanum Medium Neutral Gray (298) with Buff White (1) spot; flanks fade dorsal-to-ventral from Drab Gray (256) to Cinnamon Drab (50), to Light Buff (2) yellowishwhite; ventral surfaces Bright White to Pale Buff (1); throat with Cinnamon- or Orange-Rufous (50, 56) brown blotches; posterolateral throat (skin overlying vocal sacs) Very Dark Brown Umber (23); posterior ventrum and inguinal region transitions to Straw Yellow (53) and Orange Yellow (8) below the cloaca; ventral surfaces of distal segments of fore- and hindlimbs Pinkish Buff (3) to Buff Yellow (5); forearm, tibia, and tarsus ventral surfaces with denser aggregation of Cinnamon- to Orange-Rufous (50, 56) brown blotches medially and Jet Black (300) laterally; ventral (palmar) surfaces of hand and (plantar) surface of foot Dusky Brown (285) with Dark Pearl Gray (290) subarticular tubercles and lighter, Pale Neutral Gray (296) to Light Pearl Gray (262) terminal discs of digits.
Measurements of holotype (mm).— SVL 74.9; HL 29.2; SL 13.0; TYM 4.4; HW 29.7; FLL 16.1; THL 40.2; TL 43.7; TAR 18.9; FL 40.8; HAL 19.1; EN 7.2; IND 5.7; FANGL 4.6; FANGH 4.1.
Variation.— Summaries of univariate morphological variation in the type series are presented in Table 2 View Table 2 . We observed no apparent sexually dimorphic color variation in our large series of paratypes; similarly, although comparisons among older specimens from particular localities (CAS, FMNH, and USNM paratypes, collected over the past century) and more recent material from some of the same, but also novel localities on Mindoro (KU, and more recent USNM specimens) revealed some variation suggestive of geographically variable color variation, these initial observations could conceivably reflect circumstances of preservation and color shifts with time. As a result, we emphasize color variation across all specimens available to us (from throughout Mindoro Island): three general ground-pattern dorsal coloration types are immediately apparent, including (1) specimens with very dark brown to black dorsal surfaces, and transverse hindlimb bars, forearm blotches, and interorbital bar indistinct or barely evident (Sepia 286, Jet Black 300; e.g., KU 302095, 303432, 308303, 335866), (2) specimens with medium brown to gray dorsal surfaces (Glaucous 291 to Brownish Olive 292 or Dark Neutral Gray 299) and darker transverse hindlimb bars, forearm blotches, interorbital bar, and labial bars all evident (the holotype [PNM 9870; Fig. 6 View FIG ] and paratypes [KU 302084–88, 302100, 302105–07, 303451, 303353], plus nearly all older CAS, FMNH, and USNM paratypes and referred specimens, collected 40–100 y ago), and (3) specimens with light gray (Pale Neutral Gray 296 to Pratt’s Gray 293) dorsal body surfaces, with a dense network or reticulum of dark gray (Plumbeous 295 to Dark Neutral Gray 299; e.g., KU paratypes 302084, 302085) throughout the body, and densely congregated into hindlimb bars, forearm blotches, a darkly pigmented interorbital bar, and labial bars. Ventral body surfaces are generally white (Buff White l to Cream White 52) and transitioning posteriorly to more yellowish white (Pale Horn 11 to Light Yellow Ocher 13) in inguinal region and posterior surfaces of the thighs. Darker pattern elements situated around posterolateral surfaces of head (chin and throat lateral edges), and lateral margins of ventral limb surfaces (lateral edges of ventral forelimbs and hindlimbs) correspond to the same general dorsal coloration patterns (above), which wrap laterally onto ventral surface and sharply fade to white. Thus, the specimens with dark dorsal coloration are also the specimens with the darkest ventral pattern elements (infralabial blotches, lateral forearm ventral coloration, ventral shank color, and anteroventral thigh pattern), and the specimens with the medium brown to gray dorsal surfaces (including most paratypes and the holotype) have corresponding medium pigmentation in these same characters contributing to the ventral pattern ( Fig. 6 View FIG ). Ventral surfaces of hands and feet are surprisingly invariant and do not differ from those described in the holotype ( Fig. 6 View FIG ), and no ontogenetic color variation was evident in our series. The one ventral color pattern character that seemed to depart from the above was the concentration of pigment contributing to throat and pectoral region. Throats ranged from homogeneously medium gray (offset and darker from ventral body surface; males KU 302104, 303432, females KU 302105, 303451) to darkly spotted brown (males KU 302095, 302106, 308313, PNM 9870 [holotype], females KU 302100, 302118, 335866), to pale cream (not differing from remainder of ventral body coloration; males KU 302085, 302086, 302089, 303353, 304132, females KU 302084, 302387, 302088). In life, some specimens have more yellowish hues in lateral head surfaces, the flanks, inguinal region, ventral hindlimb and cloacal color ( Fig. 7 View FIG ); these are lost and fade to shades of cream to light gray in preservative; iris gold above and silver below pupil.
Distribution.— Limnonectes beloncioi is known only from Mindoro and Semirara Islands, central Philippines ( Fig. 1 View FIG ). Minor land-bridge and deep-water islands to the east and southwest of Mindoro have not yet been surveyed for herpetofauna; as such, the identity of any populations of Limnonectes that might eventually be discovered on Ilin, Sibay, Ambulong, Caluya, Maestre de Campo, and/or the Cuyo Islands group cannot yet be confirmed. The genetic identification of populations of Limnonectes on Busuanga, Coron, and Culion (currently considered L. acanthi [ Taylor, 1923, 1928; Inger, 1954; Brown and Alcala, 1970; Diesmos et al., 2015]; thus, we anticipate these populations will be more closely related to Palawan Island L. acanthi than to L. beloncioi ) would be of particular interest to herpetologists, biogeographers, and speciation geneticists (Esselstyn et al., 2010; Brown et al., 2013).
Etymology.— We derive the specific epithet, a patronym, to honor the Philippine army scout and freedom fighter Esteban Beloncio, who contributed substantially to the armed resistance against the Japanese WWII occupation of Mindoro Island, 1942–1945. Suggested common name ¼ Mindoro Fanged Frog.
Ecology and natural history.— The new species is commonly encountered in riparian habitats, and in the vicinity of moving water in galley forests; it has been collected from rocks, sandy shingles, and muddy river banks of streams and rivers of Mindoro Island and its land-bridge associated island, Semirara; the species also is anticipated to be present on Ilin and Ambulong Islands, which are separated from Mindoro by shallow channels. On Mindoro, the new species has been collected from just above sea level in coastal lowland habitats associated with river mouths (the type locality: Barangay Carmundo, Sitio Paypay-Ama, at the mouth of the Paypay-Ama River; the Municipality of Victoria, Barangay Loyal, Sitio Panguisan, Panguisan River), and in coastal habitats in the northern Mindoro Municipalities of Puerto Galera, Naujan, Calapan, and San Teodoro, southern Municipalities of San Jose, Bulalacao, and Roxas, and western coast Municipalities of Bongabong, Gloria, and Pinamalayan; Mindoro’s interior regions, in which the new species has been recorded, collected, or observed include the relatively undisturbed low to mid-elevation forests of the Municipalities of Sablayan ( Fig. 8 View FIG ), San Teodoro, Calapan, and Baco ( Brown and Guttman, 2002; Brown et al., 2009).
Other native species of amphibians encountered on Mindoro Island ( Diesmos et al., 2015) include Platymantis corrugatus (Ceratobatrachidae) , Fejervarya moodiei , F. vittigera , Occidozyga laevis (Dicroglossidae) , Kaloula conjuncta , K. picta (Microhylidae) , Leptobrachium mangyanorum (Megophryidae) , Pulchrana mangyanum (Ranidae) , Polypedates leucomystax , Philautus schmackeri , and Rhacophorus pardalis (Rhacophoridae) . Invasive species of frogs introduced to Mindoro include Rhinella marina (Bufonidae) , Kaloula pulchra (Microhylidae) , Hylarana erythraea (Ranidae) , and Hoplobatrachus rugulosus (Dicroglossidae) . The last of these is a voracious predator, which we have observed on numerous occasions, feeding on juveniles and metamorphs of Limnonectes .
With very little original vegetation present at lower elevations on Mindoro Island, and with the rising human population, the loss of original forest, development, and agriculture throughout most the island’s lower elevations, the loss of forest cover must be viewed as a primary threat to all native forest-associated amphibians, including L. beloncioi . Estimates of forest loss or severe alteration between 2003 and 2010, for example, range from 8–9% annually, of 53–61% cumulatively for over this 7-yr period (ELBR, pers. comm. with Center for Conservation Innovations [unpubl. data, 2020]). Despite the resilience of the new species in heavily disturbed and highly degraded riparian habitats ( Brown and Guttman, 2002; Brown et al., 2009; ELBR and RMB, pers. obs.), its ubiquitous persistence in Mindoro’s chronically degraded lowland watersheds ( Lizuka et al., 2009) suggests that poor water quality may be less of a threat to larval development in L. beloncioi than might otherwise be expected of amphibians with aquatic larval development ( Wells, 2007).
An important source of protein in the diets of many indigenous communities (including the eight ethnolinguistic Mangyan tribal groups of Mindoro; Lopez, 1976; Kikuchi, 1984), species of fanged frogs are widely hunted and consumed in alarming numbers throughout their ranges in the Philippines (RMB, CDS, and ELBR, pers. obs.). Although the new species is one of the most common species of frogs on Mindoro Island (as evinced by numbers of specimens available for this study in collections), it is heavily hunted and widely consumed by humans on Mindoro ( Schult, 1991), including the migrant Tagalog (Luzon-derived) majority in the lowlands ( Scheffers et al., 2012) and interior highlands indigenous peoples (Mangyans) who target river frogs (‘‘Palakang Ilog’’) during the rainy season (ELBR, pers. obs.). We have evaluated this species against the IUCN conservation status classification rubric ( IUCN, 2020), and find that it does not qualify for Critically Endangered (CR), Endangered (EN), Vulnerable (VU), or Near Threatened (NT) status. Limnonectes beloncioi has a geographic distribution that spans multiple islands, including the large landmass of Mindoro, and is quite abundant at all sampled localities. We therefore classify this species as Least Concern ( IUCN, 2020), but acknowledge that some other, currently undefined, threatened status may be conceivable, following consideration of unrelated, or secondary sources of information which do not bear directly on the biology of the new species ( Gonzalez et al., 2018).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Limnonectes beloncioi
| Herr, Mark W., Goyes Vallejos, Johana, Meneses, Camila G., Abraham, Robin K., Otterholt, Rayanna, Siler, Cameron D., Rico, Edmund Leo B. & Brown, Rafe M. 2021 |
Limnonectes cf. acanthi
| Evans 2003 |
Limnonectes (Limnonectes) acanthi
| Dubois 1987 |
Rana acanthi
| Taylor 1923 |
Rana macrodon blythi
| Boulenger 1920 |