
Parapodemus SCHAUB, 1938
|
publication ID |
https://doi.org/10.2478/if-2017-0021 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF8797-FFD5-FFA5-FC59-FDDFBBB9FF72 |
|
treatment provided by |
Diego |
|
scientific name |
Parapodemus SCHAUB, 1938 |
| status |
|
Genus Parapodemus SCHAUB, 1938
C o m m e n t s. The common practice of attributing all Apodemus -like fossil murids regardless of the degree of development of t7 to the genus Apodemus may appear logical if it had not been for the deep split between the recent crown groups of Apodemus s. l. into several obviously genus-rank taxa. Most of European Late Miocene and Plio-Pleistocene apodemyines could belong to the genera Sylvaemus and Karstomys . Therefore we prefer to use Parapodemus for the earliest (late Vallesian and early Turolian) and morphologically primitive forms not yet manifesting affinities to any particular group of recent apodems and conforming to the diagnosis presented by de Bruijn et al. (1999).
Parapodemus lugdunensis SCHAUB, 1938 Pl. 8, Text-fig. 12 View Text-fig
M a t e r i a l. 15 m View Text-fig 1 View Text-fig , 8 m View Text-fig 2, 3 m3 View Text-fig , 9 M View Text-fig 1 View Text-fig , 4 M View Text-fig 2, 3 M3 View Text-fig : Gaverdovsky ; 9 m 1, 6 m2, 5 m3, 8 M1, 2 M2, 1 M3: Volchaya Balka .
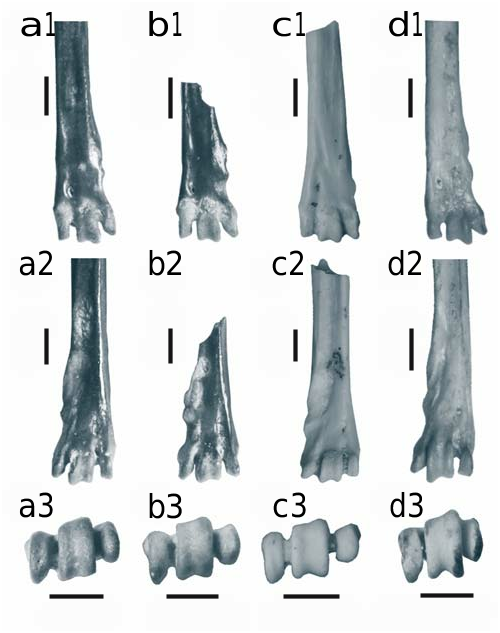
D e s c r i p t i o n. m1. The molar is composed of the tma, and usually connected anteroconid and metaconidprotoconid chevrons, and the posterior hypoconid-entoconid chevron. The anterior cusp (tma) is relatively large, elevated to the level of the anterior chevron of the anteroconid, and always present. The cusps of the labial cingulum are well developed. There are from two to four (most often three) accessory cusps anteriorly of c1. In one specimen, an accessory cusp lingual to interoanteroconid is present. The posterior heel (posteroconid) is expressed as an oval cusp. Two roots.
m2. The molar is composed of equally developed metaconid-protoconid and hypoconid-entoconid chevrons. In two specimens, the chevrons are connected by a low central ridge. The cingular cusps are variably developed. There are from 0 to 3 accessory cusps between c1 and the antero-labial cusp. Posterior heel (posteroconid) is expressed as an oval cusp, but may be reduced to a low posterior ridge closing the posterosinusid. In one specimen c1 is not expressed. Two roots.
m3. The molar is composed of the metaconid-protoconid chevron, and the posterior cusp. They may be connected by a very low lingual ridge. The antero-labial cusp is weakly developed. No conspicuous accessory cusps are present except for a minuscule swelling on the labial side of the protoconid in one specimen. Two roots.
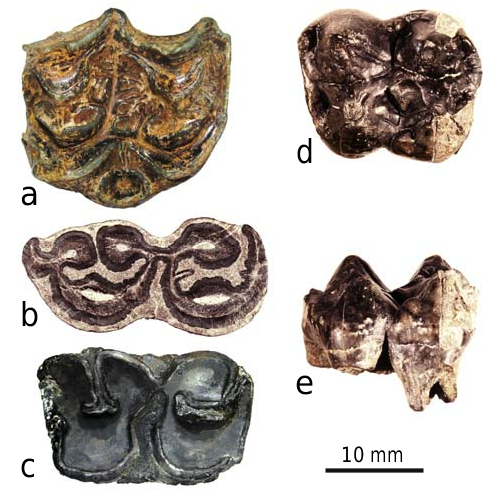
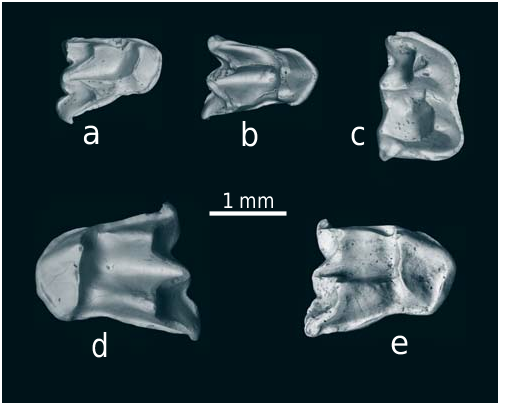
M1. The molar is composed of the anterior transverse chevron (t1-t2-t3) and posterior cusp ring with t4-t5-t6, t8-t9, and t12. The antero-lingual cusp (t1) is shifted more posteriorly relative to t2-t3 block. In some specimens t1 may connect to t2-t3 at a right angle (Pl. 8, Fig. 9) resembling the Progonomys condition. A very small t1bis is present in 2 out of 13 complete specimens. A prestile at the base of t2 is present in one molar (Pl. 8, Fig. 8). There is a posterior ridge in t3 (n = 8 out of 16). The t4 and t8 connection (Pl. 8, Figs 8–11) ranges from a direct contact without any swellings (n = 6/16), an elevated ridge sometimes turning with wear into a small swelling (n = 8/16), or a small cusp (t7) on the ridge between t4 and t8 (2/16). Cusps t6 and t9 are always connected. The postero-labial cusp t12 is well developed and may be expressed as a large circular cusp (Pl. 8, Figs 9, 10). Three roots. The lingual root may be flattened and show weak signs (longitudinal grove) of separation into two roots.
M2. The molar structure is similar to that of M1 except for the absence of t2. The cusp t1 is oval in shape with the long axis running somewhat postero-lingually. In one specimen (n = 1/5) the t1 is subdivided into two cusps (Pl. 8, Fig. 13). The cusp t3 is smaller than t1. In one specimen it is strongly reduced. The connection between t4 and t8 can appear as a high ridge (n = 3/5), or as a small t7 (2/5). Three roots (n = 3). In one specimen the lingual root is strongly flattened and slightly bifid at the apex.
M3. The molar is subtriangular. In the smaller (L = 0.87) of the two moderately used teeth, t1 is drop-shaped with a horizontally oriented long axis, t3 is missing, t4-t5-t6 broadly fused, t8 communicates with t6 and with a ridge-like connection with t4 (Pl. 8, Fig. 15). The larger molar (L = 1.1) has a much more complex structure with cusplet-like t3, large crescent obliquely oriented t1, discernable t9, low ridge-like connection between t4 and t8, and even a notch-like t12 on the posterolabial side of t8 (Pl. 8, Fig. 16). Three roots.
C o m m e n t s. The Caucasian form shows slightly larger size values in the upper part of the observed variation range and has a higher percentage of weak t7 formation in M1 (but less than 50%) than in P. lugdunensis from the late Vallesian and early Turolian sites of Central Europe (Schernham, Kohfidisch, Eichkogel) and Iberia (Los Aguanaces 3, Crevillente 2 and 4B) ( Daxner-Höck 1977, van de Weerd 1976, van Dam 1997, Martín Suárez and Freudenthal 1993, Wöger 2011, Daxner-Höck and Höck 2015) but lower than in the Anatolian site of Düzyayla (de Bruijn et al. 1999) attributed to the middle Turolian. This can be taken as an indication of a late early Turolian age for the Caucasian form. Nesin (2011) described a small species Apodemus alae diagnosed on the presence of t 7 in less than 50% of specimens from the early and middle Turolian faunas in south Ukraine. The large sample from the type locality, the middle Turolian fauna of Cherevichnoe 3, shows t 7 in approximately 20–30% of M1–2 ( Nesin 2013). This species is likely to be a younger synonym of P. lugdunensis .
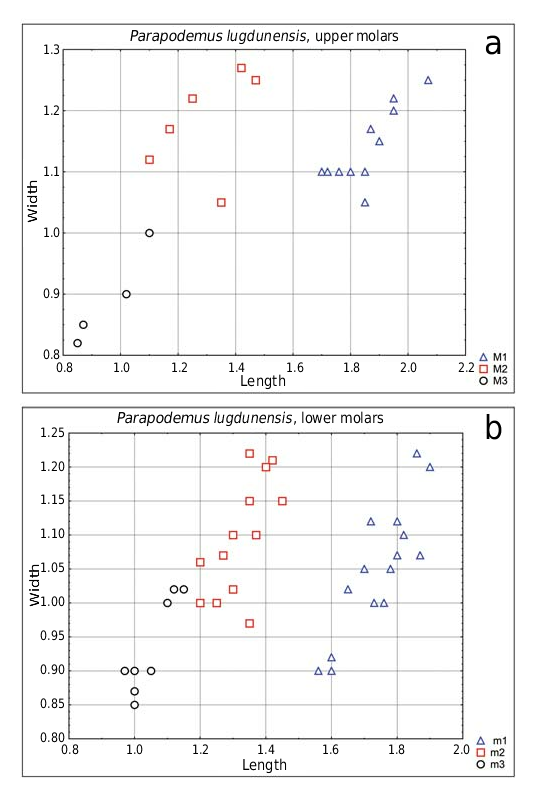
The size distribution is quite continuous in most molar types ( Text-fig. 12 View Text-fig ), except for M3. In this latter case the sample size (n = 4) is very low therefore impeding any reliable conclusions. The very different morphology of two moderately worn M3 may indicate a considerable morphotype variation within a single species (as illustrated for P. lugdunensis from Kohfidisch by Wöger 2011) or alternatively, a co-occurrence of a smaller and larger species. This hypothetical large form would then be represented by only a few specimens. A possible admixture of a larger form may also be signalled by somewhat high CV values for some molar types ( Tab. 4). At the same time the largest M1 (Pl. 8, Fig. 8) shows outlines similar to that of smaller specimens (Pl. 8, Figs 9–12) and does not show any variation towards larger W/L values in M1 known for P. barbarae , the larger species commonly co-occurring with P. lugdunensis in the middle Turolian of Spain (Los Mansuetos) and Ukraine (Cherevichnoe 3). Only a larger sample can shed light on this issue.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.