
Kaiparathina monticola, Herbert, 2024
|
publication ID |
https://doi.org/ 10.5852/ejt.2024.923.2445 |
|
publication LSID |
lsid:zoobank.org:pub:AD68CEDD-2F2D-4010-BE7A-1B1AE9E4A0F3 |
|
DOI |
https://doi.org/10.5281/zenodo.10818420 |
|
persistent identifier |
https://treatment.plazi.org/id/C4A503AD-4E0D-4DF7-8A82-ECF326086C36 |
|
taxon LSID |
lsid:zoobank.org:act:C4A503AD-4E0D-4DF7-8A82-ECF326086C36 |
|
treatment provided by |
Plazi |
|
scientific name |
Kaiparathina monticola |
| status |
sp. nov. |
Kaiparathina monticola View in CoL sp. nov.
urn:lsid:zoobank.org:act:C4A503AD-4E0D-4DF7-8A82-ECF326086C36
Figs 38C–E View Fig , 39–40 View Fig View Fig
Diagnosis
Shell elevated-turbiniform, height up to 8.0 mm; whorls strongly convex, periphery often weakly angular, well below mid-whorl; early teleoconch whorls with fine, close-set, spiral threads, but shell smooth and glossy thereafter; anomphalous; 1–3 indistinct spiral cords adjacent to umbilical region; columella noticeably thickened, reflected over umbilical region; shell pale corneous brown, variously marked with white blotches often in two spiral bands, occasionally merging into zig-zag axial flames; protoconch white, its tip sometimes tinged maroon.
Etymology
From the Latin “ mons ” – “a mountain” and “ -cola ” – “a dweller”; reflecting the seamount habitat of the species.
Material examined
Holotype
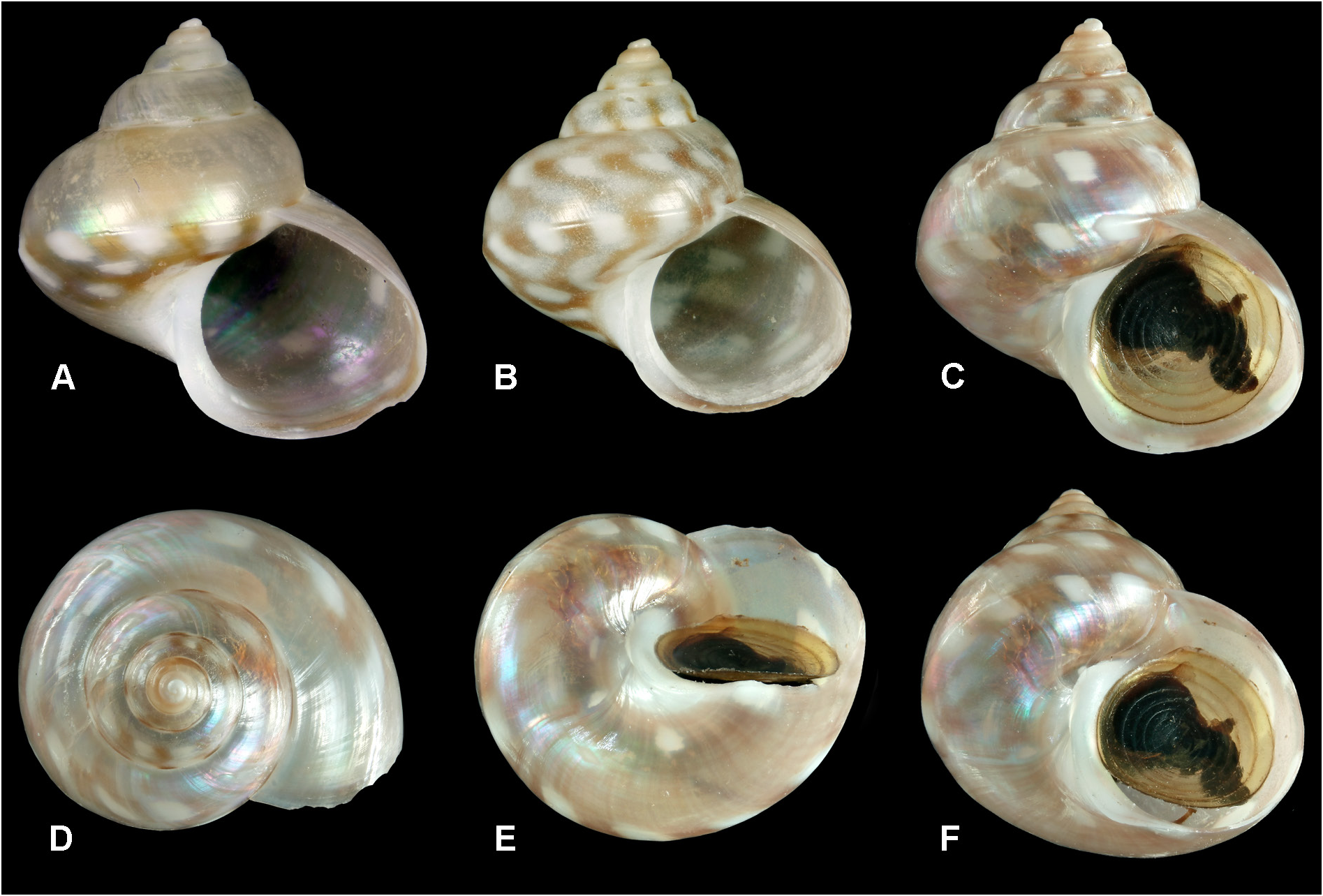
WALTERS SHOAL • living specimen; slopes, stn DW4881; 33°16′ S, 43°50′ E; depth 377–382 m; 2 May 2017; DNA tissue sample, photo alive ( Fig. 38C–E View Fig ); MNHN-IM-2013-67168 .
GoogleMapsParatypes
WALTERS SHOAL – slopes • 1 living specimen; same collection data as for holotype; DNA tissue sample; MNHN-IM-2013-67167 • 4 specimens, living; same collection data as for holotype; MNHN- IM-2000-35726 GoogleMaps • 1 living specimen; stn DW4890; 33°09′ S, 43°59′ E; depth 492–588 m; 4 May 2017; DNA tissue sample; MNHN-IM-2013-67258 GoogleMaps • 1 living specimen; stn DW4890; 33°09′ S, 43°59′ E; depth 492–588 m; 4 May 2017; DNA tissue sample; MNHN-IM-2013-67259 GoogleMaps • 1 living specimen; stn DW4897; 33°09′ S, 43°59′ E; depth 490–584 m; 5 May 2017; DNA tissue sample; MNHN- IM-2013-67260 GoogleMaps • 5 specimens, living; stn DW4897; 33°09′ S, 43°59′ E; depth 490–584 m; 5 May 2017; radula SEM prep.; MNHN-IM-2000-35727 GoogleMaps • 2 specimens, living; stn DW4898; 33°09′ S, 44°01′ E; depth 652–668 m; 6 May 2017; MNHN-IM-2000-35728 . GoogleMaps
GoogleMapsOther material
WALTERS SHOAL – slopes • 40 + empty shells; same collection data as for holotype; MNHN GoogleMaps • 1 empty shell; stn DW4885; 33°17′ S, 43°55′ E; depth 272–380 m; 3 May 2017; MNHN GoogleMaps • 6 empty shells; stn DW4886; 33°17′ S, 43°56′ E; depth 573–582 m; 3 May 2017; MNHN GoogleMaps • 2 empty shells; stn DW4887; 33°17′ S, 43°57′ E; depth 599–640 m; 3 May 2017; MNHN GoogleMaps • 11 empty shells; stn DW4890; 33°09′ S, 43°59′ E; depth 492–588 m; 4 May 2017; MNHN GoogleMaps • 15 empty shells; stn DW4897; 33°09′ S, 43°59′ E; depth 490–584 m; 5 May 2017; MNHN GoogleMaps .
Description
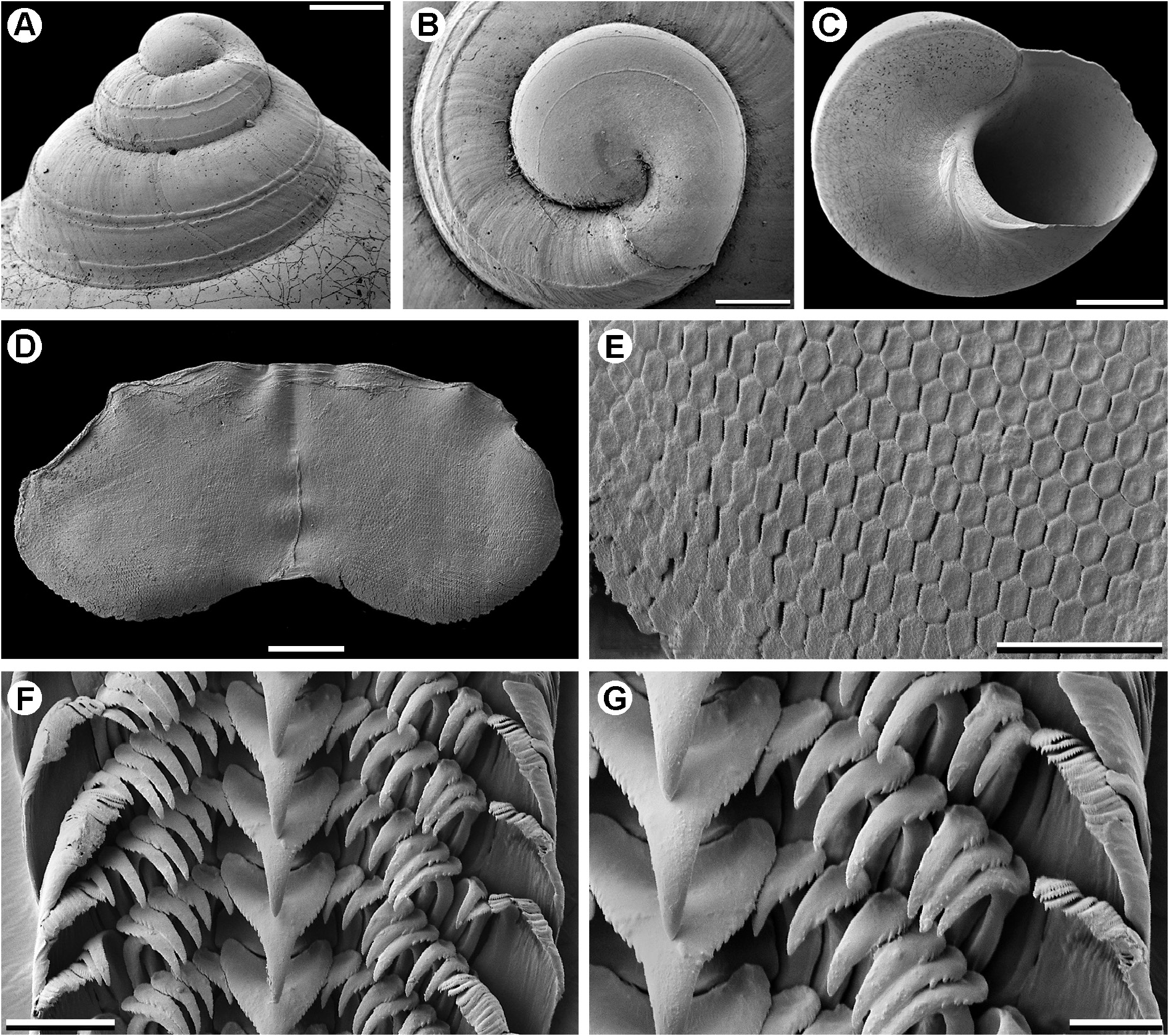
SHELL. Relatively large for the genus (height up to 8.0 mm), elevated-turbiniform, slightly higher than wide; spire prominent with strongly convex whorls; periphery often weakly angular, well below mid-whorl; suture distinctly indented; sculpture weak, shell essentially smooth and glossy; base less strongly convex, umbilicus lacking; teleoconch of up to 5.0 whorls. First two teleoconch whorls with pair of fine, close-set, spiral threads midway between sutures ( Fig. 40A View Fig ), second weaker pair below this, lower one of which close to or level with abapical suture; upper pair each developing shallow adapical groove during second whorl; spiral sculpture evanescing near start of third whorl and later whorls with at most traces of spiral sculpture, though in live-taken specimens spiral patterning in underlying nacre visible by transparency; axial sculpture of weak growth-lines and close-set, microscopic, collabral threads; juvenile and subadult shells with more angular periphery, delineated by low spiral cord; insertion of suture level with cord; cord weak or absent on much of last whorl in adult specimens; base sculptured as above, often with 1–3 indistinct spiral cords adjacent to umbilical region; growth-lines sigmoidal below periphery and somewhat stronger medially; umbilicus lacking. Aperture subcircular; columella noticeably thickened, reflected over umbilical region; outer lip thin, concave below periphery, convex near columella, basal lip thus sinuous; interior nacreous, lacking labral ornamentation.
PROTOCONCH ( Fig. 40A–B View Fig ). Globose, somewhat elevated, comprising approx. 1.25 whorls, diameter 350–375 μm; tip of apex roundly angled, terminal lip convex, lacking terminal or subterminal varix; surface smooth save for three fine, widely spaced, subspiral threads, lowest one close to abapical suture, a fourth one delineating tip of apex.
COLOUR. Protoconch white, its tip sometimes tinged maroon; teleoconch with pale corneous brown ground colour, translucent in live-taken specimens, with pink/green iridescence of underlying nacre showing through. Colour pattern variable; commonly with two spiral rows of white blotches ( Fig. 39C View Fig ), one at periphery, the other between this and adapical suture, sometimes a third row near middle of base; intervals between white blotches somewhat darker brown, particularly at periphery, on spire whorls this creates a thin supra-sutural line of alternating white/brown dashes; often also with thin subsutural white line; columella and medial area of base white; variation in white and brown markings considerable, the latter sometimes merging into zig-zag axial flames ( Fig. 39B View Fig ); some specimens with broad whitish subsutural band, others with only alternating light and dark peripheral blotches or with blotches absent on spire whorls or present only on spire whorls.
DIMENSIONS. Holotype, height 6.9 mm, diameter 6.8 mm; largest specimen, height 8.0 mm, diameter 7.6 mm.
OPERCULUM. Corneous, thin, light yellowish-brown, tightly multispiral with a short growing margin.
JAW ( Fig. 40D–E View Fig ). Oval-quadrate, lateral portions rounded and dipping anteriorly, posterior edge thus weakly convex and anterior edge weakly concave; surface composed of minute, elongate hexagonal units (occasionally pentagonal).
RADULA ( Fig. 40F–G View Fig ). Formula ∞+7+1+7+∞, with ± 35 transverse rows of teeth; rachidian tooth very large and robust, its cusp basally trigonal but with convex sides; cutting edge dominated by large, narrowly pointed central denticle; basal part of cusp with finer lateral denticles; anterior margin of cusp concave, abutting convex posterior edge of base-plate of next anterior tooth. Lateral teeth slender, scythe-like, cusp elongate and slightly curved, its outer base flanged and bearing small denticles; shaft face with thickened pilaster; laterals 3–5 largest. Marginal teeth numerous, progressively reducing in size toward edge of radula, shafts of all but innermost one seemingly fused; cusps of inner marginals narrowly spathulate, outer edge minutely denticulate.
EXTERNAL ANATOMY ( Fig. 38C–E View Fig ). As described for Kaiparathina boucheti ( Marshall 1993) . Animal for the most part milky-white; head and neck tinged with pale apricot-orange; cephalic tentacles well developed, micro-papillate and with a faint orange-brown mid-dorsal line; eyes prominent, pigmented and with open aperture; anterolateral structures pale, coarsely pustulose (arrowed in Fig. 38E View Fig ), lying beneath non-digitate neck lobes; three unpigmented, micro-papillate epipodial tentacles on each side, each with basal epipodial sense organ; sides of foot beneath epipodium orange-brown, colour progressively deepening ventrally, but then ceasing abruptly, lower portion comprising uniformly pale band, encircled by thinner orange-brown band (evanescing prior to propodial horns) and finally pale peripodial fringe; colour pattern continues onto metapodium, which has white mid-line band at extreme posterior ( Fig. 38C View Fig ).
Distribution
Known only from the slopes of Walters Shoal, at depths of 380–652 m (live-taken material 382–652 m); dredged on substrata of coarse sand and fine gravel, with ophiuroids and solitary corals.
Remarks
In terms of its relatively large size, Kaiparathina monticola sp. nov. resembles K. coriolis Marshall, 1993 and K. boucheti Marshall, 1993 , both from the New Caledonian region. Kaiparathina coriolis has more strongly convex spire whorls, retains an umbilical chink at maturity and has a different coloration. In addition, the terminal cusp of the rachidian tooth is proportionately much smaller than it is in K. monticola . Kaiparathina boucheti has stronger spiral sculpture on the apical whorls, is more uniform in colour and has a reddish-brown protoconch; the rachidian tooth also has a more distinctly triangular cusp. Perhaps the most similar species is K. daedala Marshall, 1993 from off Réunion and this is also the species most geographically proximate to Walters Shoal. Like the present species, K. daedala has a narrow supra-sutural band of reddish-brown marks (also present in K. coriolis ) and a narrow white subsutural line. Elsewhere, the colour pattern in the only two available specimens is variable ( Marshall 1993), but neither exhibit the spiral lines of white spots or zig-zag brown axial flames commonly seen in K. monticola . In addition, a distinctive feature of K. daedala is the presence of numerous spiral threads on the inner two-thirds of the base. Specimens of K. monticola of a similar size (4–5 mm) may have 1–2 such spiral threads, but in most individuals they are absent ( Fig. 40C View Fig ). The radula described for K. daedala has only five pairs of lateral teeth in each row (vs seven in K. monticola ), but this difference is undoubtedly size-related and reflects the fact that the type material of K. daedala was probably subadult, the number of lateral teeth increasing progressively through in-column transformation of marginal teeth during ontogeny ( Marshall 1993). There are, nonetheless, differences in the shape of the radula teeth, the terminal denticle of the rachidian and lateral teeth being proportionately longer in K. monticola , whereas the subterminal denticles on these teeth are coarser and more distinct in K. daedala . Furthermore, in K. monticola the cusp of the rachidian is laterally more convex at its base than the more evenly trigonal rachidian cusp of K. daedala . Although likely to be closely related to K. daedala , these differences in coloration, basal sculpture and radula tooth morphology justify the separation of the present material as a distinct species.
The shell of K. monticola sp. nov. superficially resembles that of some brown-spotted species of Bathymophila ( Williams et al. 2020: fig. 7g), which Williams et al. (2022) have shown belong to an undescribed Bathymophila -like genus. These differ from K. monticola in having a patent umbilicus with a heavy flange around the umbilicus, and a less acutely pointed apex.
Mention should also be made of Cantharidus nolfi Poppe, Tagaro & Dekker, 2006 from the Philippines. Although not known anatomically, I believe the shell of this species exhibits all the features characteristic of Kaiparathina and I propose that it be transferred thereto. It is of similar size as K. monticola sp. nov., but differs in having a colour pattern that typically includes fine, close-set and strongly oblique gold or brown lines.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Vetigastropoda |
|
Order |
|
|
SuperFamily |
Trochoidea |
|
Family |
|
|
SubFamily |
Kaiparathininae |
|
Genus |