
Diognetus cheimon, Yasunaga & Schwartz & Chérot, 2023
|
publication ID |
https://doi.org/ 10.37520/aemnp.2023.001 |
|
publication LSID |
lsid:zoobank.org:pub:3F2C90B1-6EA1-4B38-A218-C314D09F6E00 |
|
DOI |
https://doi.org/10.5281/zenodo.7887774 |
|
persistent identifier |
https://treatment.plazi.org/id/6A287DC8-3F9A-43B5-BAE3-7FA9F59343D1 |
|
taxon LSID |
lsid:zoobank.org:act:6A287DC8-3F9A-43B5-BAE3-7FA9F59343D1 |
|
treatment provided by |
Felipe |
|
scientific name |
Diognetus cheimon |
| status |
sp. nov. |
Diognetus cheimon sp. nov.
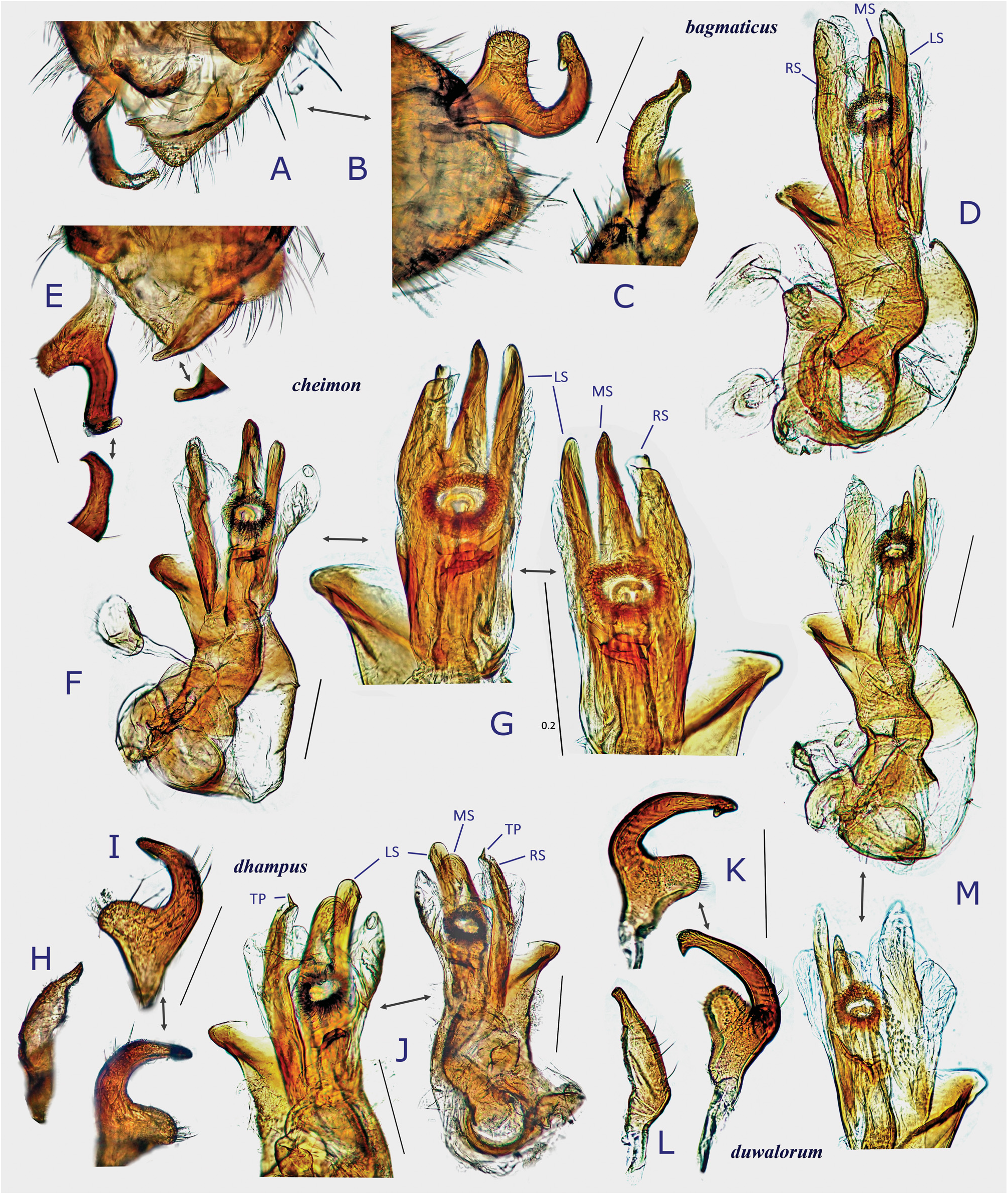
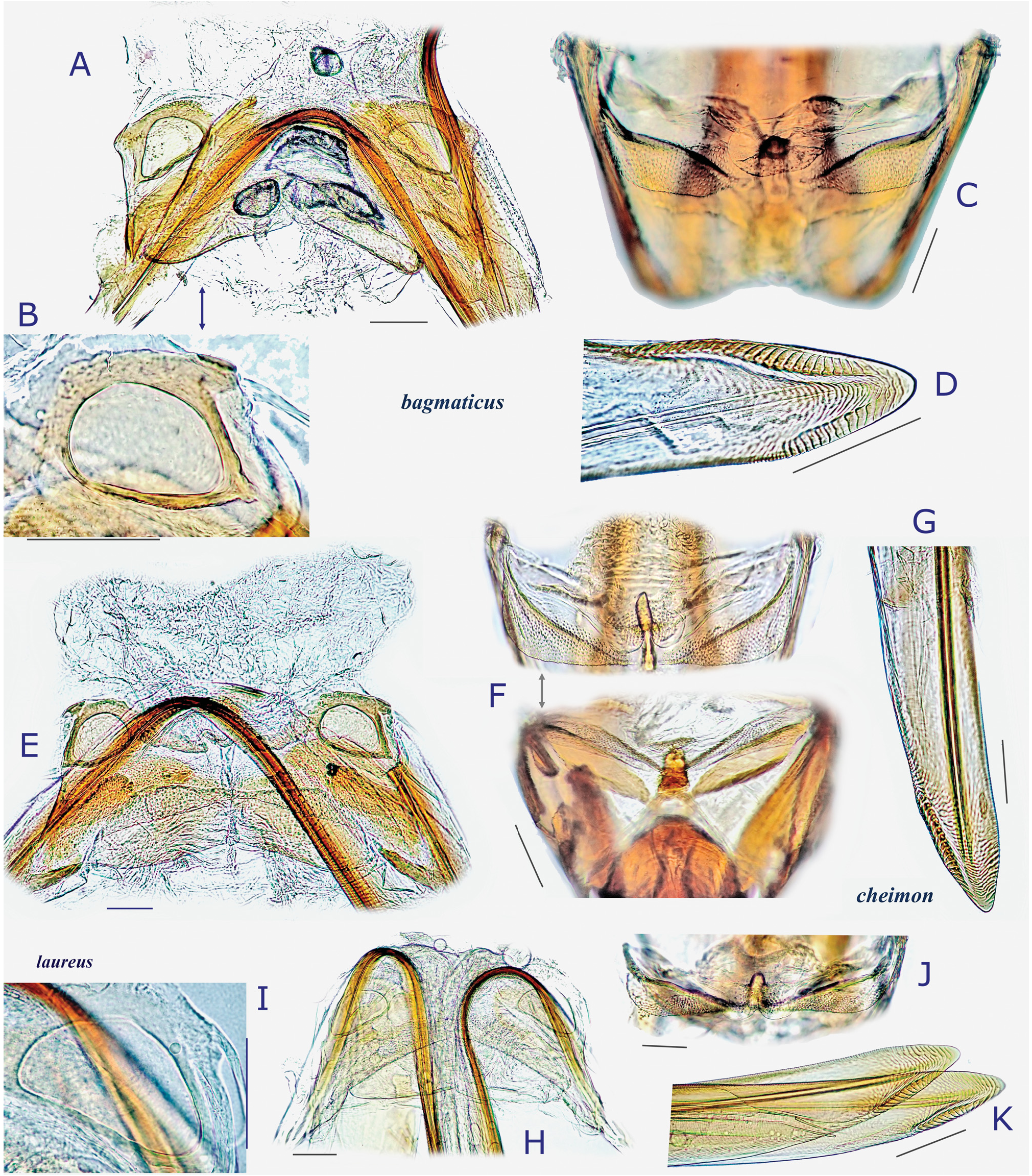
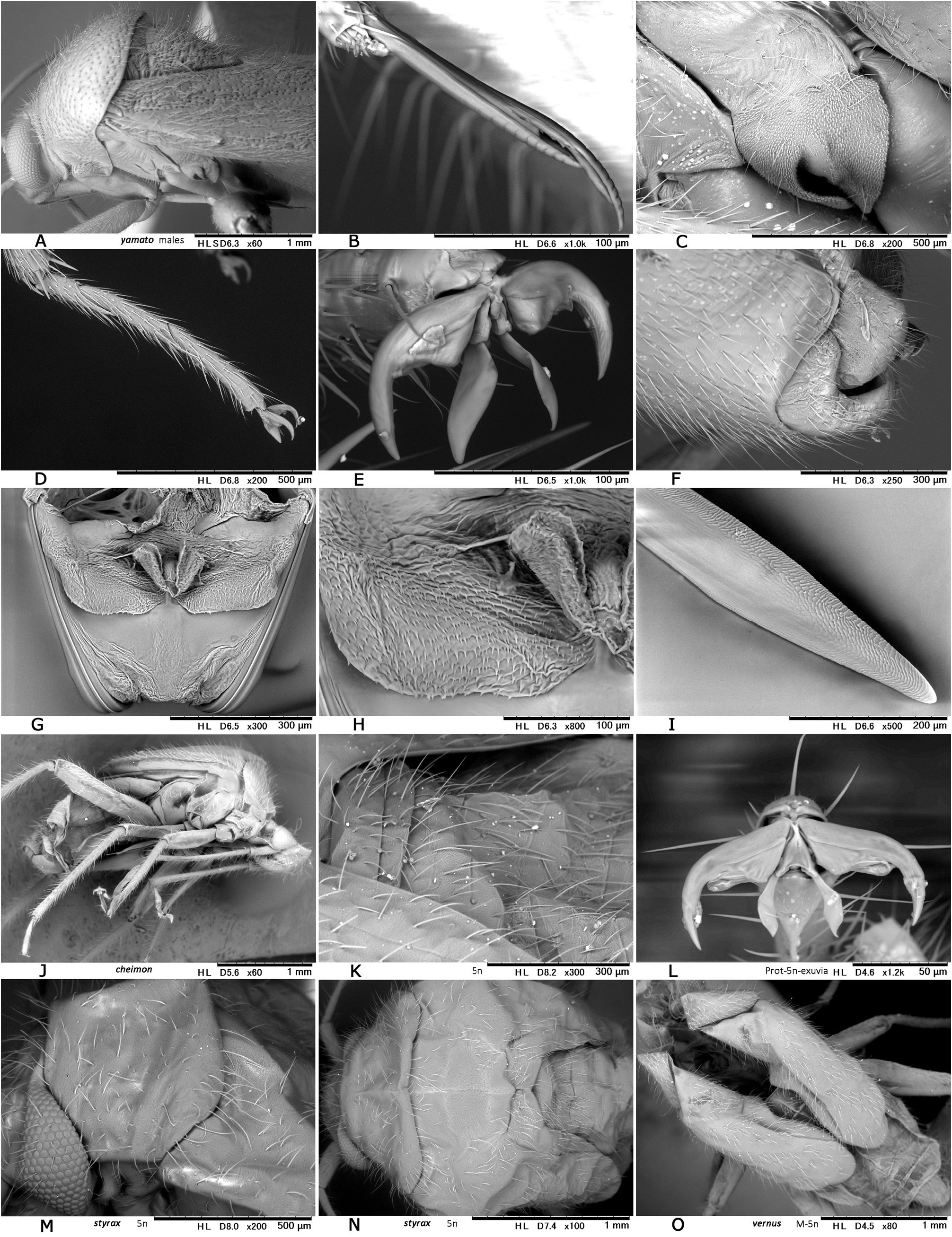
( Figs 4 View Fig , 5A–H View Fig , 8E− G View Fig , 10E− G View Fig , 23 View Fig , 26 View Fig , 36J− L View Fig )
Type material. HOLOTYPE: J, JAPAN: KYUSHU: Nagasaki Pref., Nagasaki City, Kurosaki-Nagata Marsh (biotope), 32.829185, 129.721365, sweeping flowers of Eurya emarginata , 13 Dec 2020, T. Yasunaga ( AMNH _ PBI 00380745 ) ( AMNH) GoogleMaps . PARATYPES: JAPAN: HONSHU: Chiba Pref., Matsudo City, 21 Century Forest Park, 35.807500, 139.938333, 9 Sep 2012, N. Muro ( TYCN). KYUSHU: Nagasaki Pref.: Nagasaki City, Iojima Island (lighthouse-park), 32.714177, 129.761155,sweeping flowers and fruits of Eurya emarginata , 27 Dec 2019, T. Yasunaga & K. Tanaka, 3 JJ 2 ♀♀ [1 J – 5th instar immature form reared then emerging on Jan 1, 2020] ( TYCN); Nagasaki City, Kabashima Island, 32.564877, 129.780177, sweeping flowers and fruits of Eurya japonica , 27 Dec 2019, T. Yasunaga, 2 JJ and 1 J [5th instar immature form reared then emerging on Dec 30, 2019] ( TYCN); Nagasaki City, Kurosaki-Nagata Marsh (biotope), 32.829185, 129.721365, sweeping flowers and fruits of Eurya emarginata , 15 Dec 2019, T. Yasunaga, 3 ♀♀ and 1 J [5th instar immature form reared then emerging on Dec 17, 2019, dead on Jan 2, 2020] ( TYCN); same locality and plant, 13 Dec 2020, T.Yasunaga, 3 ♀♀ ( TYCN) and 1 ♀ [5th instar immature form reared then emerging on Dec 15, 2020] ( TYCN); Nagasaki City, Sotome, Higashi-Shitsu, 32.835325, 129.702280, sweeping flowers of Eurya emarginata , 13 Dec 2020, T.Yasunaga, 1♀ ( TYCN); Nagasaki City,Konoura-Ohgiyama,Konoura Dam, 32.888, 129.722, UV lighting, 20 Mar 2016, T. Nozaki, 1 J ( TYCN), 1♀ ( WCF); same data, except for date 27 May 2016, 1 ♀ ( TYCN); Nagasaki City, Tateyama, Mt. Konpira, sweeping flower buds of Eurya japonica , 32.764888, 129.879500, 13 Nov 2021, T. Yasunaga, 1 ♀ ( TYCN); same data, except for date 24 Dec 2021, 1 J 2 ♀♀ ( TYCN); Nagasaki City, Takenokubo, Nagasaki West High School, 32.765877, 129.859789, inflorescence of Osmanthus heterophyllus , 9 Dec 2021, T. Yasunaga, 1 J 2 ♀♀ ( TYCN); Nagasaki City, Nameshi 2-24, 32.809253, 129.841743, on Eurya emarginata , 6 Dec 2020, T. Yasunaga, 1 J ( TYCN); Nagasaki City, Taira (Azekari, Fishery Port), 32.807615, 129.770780, sweeping flowers and fruits of Eurya emarginata , 6 Dec 2020, T. Yasunaga, 2 JJ 1 ♀ ( TYCN) and 1J 1 ♀ [5th instar immature form reared then emerging on Dec 8–10, 2020] ( TYCN); same locality and plant, 13 Dec 2020, 2 ♀♀ [5th instar immature form reared then emerging on Dec 18, 2020] ( TYCN); same locality and plant, 29 Nov 2021, 2 JJ 1 ♀ [4–5th instar immature form reared then emerging on Dec 1–7, 2021] ( TYCN); Nagasaki City, Kinkai-Tonehara, 32.900888, 129.795522, sweeping buds of Eurya japonica , 21 Jan 2020, T. Yasunaga, 1 ♀ [freshly emerged and dead on May 7, 2020] ( TYCN); Nishisonogi-gun,Togitsu-Nagayo, Sakino Nature Park, 32.854271, 129.854646, sweeping buds of Eurya japonica , 20 Dec 2020, T. Yasunaga, 1 J 1 ♀ and 1 ♀ [5th instar immature form reared then emerging on Dec 21, 2020] ( TYCN); same data, 1 J 1 ♀ [5th instar immature form reared then emerging on Dec 25 (J) / Dec 28 (♀), 2020 and dead on Apr 28, 2021] ( TYCN); Omura City, Kushima, 32.897077, 129.952977, sweeping flowers of Eurya emarginata , 15 Jan 2020, T.Yasunaga, 1J [5th instar immature form reared then emerging on Jan 21, 2020] ( TYCN); same locality and plant, 9 Dec 2020, T. Yasunaga, 2 ♀♀ [5th instar immature form reared then emerging on Dec 15–18, 2020] ( TYCN); same locality and plant, 23 Dec 2020, T. Yasunaga, 1 J ( TYCN); Saikai City,Ohseto Town, Fukushima,Shirikusari Beach, 32.925066, 129.640533 sweeping fruits of Eurya emarginata , 2 Jan 2020, T. M. & H. Yasunaga, 1 J; Saikai City, Oshima, Mt. Yuridake, 33.040, 129.608, UV lighting, 4 May 2016, T. Nozaki ( TYCN). SHIKOKU: Kochi Pref., Tosa-Shimizu City, Cape Ashizuri, 32.73, 133.00, UV lighting, 10 Apr 1999, T. Befu, 1 J 1 ♀ ( TYCN); Kochi Pref., Cape Muroto, Muroto Skyline Road [Rt 203], 33.25232, 134.17835, sweeping Eurya emarginata , 18 Dec 2019, M. Takai, 15 JJ 24 ♀♀ ( AMNH, CNC, NIAES, NMPC, TYCN); same locality, beating Eurya emarginata , 9 Dec 2000, T. Befu ( TYCN); Kochi Pref., Tosa City, Mt. Yokose-yama, 33.4693, 133.4377, UV lighting, 27 March 2000, T. Befu, 1 ♀ ( TYCN); Kochi Pref., Sukumo City, Nakasuji Dam, 32.9282, 132.8137, UV lighting, T. Befu, 1 ♀ ( TYCN).
Additional material examined. JAPAN: KYUSHU: Kumamoto Pref., Amakusa City, Tsuruha-yama Park, 32.15, 130.04, UV lighting, 18 Oct 2015, T. Nozaki, 3 JJ 2 ♀♀ (all teneral specimens, WCF).
Description. Body elongate ovoid, moderate to large in size, 4.8–6.1 mm. COLORATION: Dorsum varying from reddish-brown ( Fig. 4B View Fig ) to chocolate brown ( Fig. 4A View Fig ), usually with mottled pattern ( Figs 4A− C View Fig ). Antennae pale reddish-brown; apex of segment II, segment III (except for pale extreme base) and segment IV dark brown. Labium pale brown, partly tinged with red. Pronotum reddish-brown, with calli and posterior half sometimes darkened ( Fig. 4A View Fig ) and posterior margin narrowly pale; pleura broadly fuscous; scent efferent system creamy yellow. Hemelytron reddish-brown, usually speckled with dark maculae; apices of exocorium (embolium) and cuneus narrowly pale; membrane smoky brown, with yellowish veins. Coxae and legs yellowish brown; mesofemur with two faint dark rings subapically; apical 1/3 of metafemur dark brown, with two pale rings subapically. Ventral surface of abdomen pale brown (somewhat greenish when alive), with more or less darkened lateral margins ( Figs 4E− F View Fig ). SURFACE AND VESTITURE: As in generic diagnosis; dorsal surface weakly shining; scutellum transversely rugose; hemelytron rather matte. STRUCTURE: Vertex with a basal transverse carina weakly arched. Antennal segment I about as thick as pronotal collar, longer than segment IV. Labium slightly exceeding apex of mesocoxa but not reaching apex of metacoxa ( Figs 4E− F View Fig ). Scutellum rather flat, shallowly and sparsely punctate, with apex weakly produced ( Fig. 26D View Fig ). Metathoracic scent efferent system as in Fig. 26C View Fig . Metatarsomere I short, about half as long as III; pretarsal structure as in Figs 26F− G View Fig ; parempodia rather long. MALE GENITALIA ( Figs 8E− G View Fig 26H–K View Fig ): Left paramere with squared, developed sensory lobe and rather broad hypophysis that is hooked at apex ( Figs 8E View Fig , 26J View Fig ). Vesica with short, branched MS and smooth LS, lacking noticeable TP ( Figs 8G View Fig , 26K View Fig ). FEMALE GENITALIA ( Figs 10E− G View Fig , 26L− P View Fig ): Sclerotized ring with thickened anterior rim ( Figs 10E View Fig , 26L View Fig ); posterior wall ( Figs 10F View Fig , 26N− P View Fig ) with narrow dorsal structure and moderate-sized interramal lobe.
Measurements. See Table 1.
Differential diagnosis. Recognized by its relatively elongate, moderate to large body; short labium slightly surpassing apex of mesocoxa; generally long wings; and shape of the male and female genitalic structures described above. Most similar in external appearance to D. bagmaticus sp. nov.; distinguished by long antennal segment IV that is obviously longer than segment I, subapically constricted sensory lobe of left paramere ( Figs 8E View Fig , 26J View Fig ), short right paramere ( Fig. 26I View Fig ); vesica with elongate MS; and wider interramal lobe with comb-like scaly microstructures laterally ( Fig. 26P View Fig ).
The final instar nymph is recognized by the following characters (cf. Figs 5A–D, E View Fig ): Body ovoid, relatively large, 3.6–3.9 mm in length; basic coloration varying from pale greenish yellow ( Fig. 5A View Fig ) to reddish-brown ( Figs 5B, E View Fig ); dorsum sometimes with mottled pattern, with densely distributed, simple, semierect setae and rather sparsely distributed, dark, stiff, upright setae ( Figs 36J–K View Fig ); antenna and legs pale greenish yellow to reddish-brown, partly with small, dark spots; abdomen with reddish spots; scent gland opening on abdominal tergum III with semicircular spot; and anal ridge with 4 dark spots ( Fig. 5D View Fig ).
Measurements. See Table 1.
Etymology. From Greek, cheimon (winter), referring to its particular emergence in December and January; most of specimens were collected during this cold season ( Table 2). Noun in apposition.
Biology. The present new species was confirmed to be associated with Pentaphylacaceae evergreen broadleaf trees, Eurya emarginata (Thunb.) Makino and E. japonica Thunb. This winter-emerging mirid was predominantly found in December and January, when the inflorescences and fruits of the host trees were abundant ( Figs 1A− C View Fig ). Within recent years, the mirid has frequently been found in urbanized zones and residential areas of southwestern Japan, as Eurya emarginata is now planted everywhere for landscaping ( Fig. 1B View Fig ) and for house hedge ( Fig. 1C View Fig ). The mirid species appears to expand its habitat to artificial environment, utilizing such available host plants. On the other hand, Eurya japonica is also common in southern Japan and sometimes used for traditional religious (Shinto) ceremonies.
A univoltine or bivoltine life cycle is assumed for Diognetus cheimon sp. nov. (cf. Table 2). The first annual generation appears to emerge in autumn (September− October) and the second generation (late instar immature forms and adult) could hibernate. In Nagasaki area, western Kyushu, the adults emerging in winter (December− January) survived until April or May when reared carefully with the host leaves and fermented milk beverage (Yasunaga, pers. observ.). Like as Diognetus vernus sp. nov. (see below), the adults of D. cheimon sp. nov. were found to have long life span (107–124 days in J / 121 days in ♀). Several adults were recently found on inflorescence of a (Japanese) holly tree, Osmanthus heterophyllus (G.Don) P.S.Green (Oleaceae) , but no immature forms were confirmed.
Distribution. Japan (Honshu, Shikoku, Kyushu); habitats are restricted to warm temperate climatic zones where the breeding hosts grow.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.