
Chrysemys corniculata, Jasinski, 2023
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlac084 |
|
publication LSID |
lsid:zoobank.org:pub:17B5CEBF-CF15-4254-9725-1C55476A0147 |
|
DOI |
https://doi.org/10.5281/zenodo.7924775 |
|
persistent identifier |
https://treatment.plazi.org/id/A1118AAC-7E71-4AD9-A130-23CB40A339B4 |
|
taxon LSID |
lsid:zoobank.org:act:A1118AAC-7E71-4AD9-A130-23CB40A339B4 |
|
treatment provided by |
Plazi |
|
scientific name |
Chrysemys corniculata |
| status |
sp. nov. |
CHRYSEMYS CORNICULATA SP. NOV.
( FIGS 2–3 View Figure 2 View Figure 3 , 5–7 View Figure 5 View Figure 6 View Figure 7 )
Zoobank registration: u r n: l s i d: z o o b a n k. org:act: A1118AAC-7E71-4AD9-A130-23CB40A339B4.
Holotype specimen: ETMNH-12491 , complete carapace and nearly complete plastron, missing only small portions near the left and right bridges and the posterior-most rim of the plastron; set of incomplete lower jaws including the left and right dentaries; proximal left coracoid fragment; humerus fragment; complete right fibula; and numerous other indeterminate shell and bone fragments.
Paratypes: ETMNH-503 , complete, but partially crushed, carapace and plastron ; ETMNH-3559 , nearly complete carapace (missing only portions of right peripherals 2–5 and 8), incomplete plastron including both epiplastra, entoplastron, fragment of right hyoplastron, nearly complete right xiphiplastron, and nearly complete hyo, hypo and xiphiplastra, nearly complete left and right quadrates, complete right radius, two carpals, three complete to nearly complete manual phalanges, three manual unguals, a proximal left femur fragment, and numerous other indeterminate and unidentified shell and bone fragments ; ETMNH-3561 , incomplete carapace and nearly complete plastron, with the latter missing portions of the right hyoplastron and both bridges, humerus fragment, and numerous other indeterminate and unidentified shell and bone fragments ; ETMNH-20544 , nearly complete carapace and plastron missing only a few small fragments including those from right peripheral 2, left costal 3 and the left hyoplastron near the bridge, nearly complete left maxilla, nearly complete cervical vertebra 7, incomplete right and fragmentary left humeri; complete? manual ungual, fragmentary right ischium, and numerous other indeterminate and unidentified shell and bone fragments ; ETMNH-20609 , nearly complete shell (ontogenetically younger individual) missing portions of neural 1, left costals 1–2 and 4–5, right costals 1 and 5, all of left peripheral 8, and portions of the left hyoand right xiphiplastron, there are other places on the shell where small fragments are missing as the shell was pieced back together, and it includes numerous other indeterminate shell fragments not attached to the major preserved portion of shell .
Referred specimens: ETMNH-4, right peripheral? 9 fragment; ETMNH-293, nearly complete right hyoplastron; ETMNH-297, nearly complete nuchal; ETMNH-301, left peripherals 7–8; ETMNH-3467, complete left epi- and hyoplastron; ETMNH-3527, left costal 6; ETMNH-3535, left peripheral 3; ETMNH-3544, nearly complete left epiplastron; ETMNH-3553, right peripheral 10; ETMNH-3567, incomplete left hypoplastron; ETMNH-3780, complete right xiphiplastron; ETMNH-7624, left peripheral 7; ETMNH-7631, nearly complete right costal 1 and indeterminate fragments; ETMNH-7632, articulated and nearly complete right hypoplastron and complete right xiphiplastron; ETMNH-7635, nearly complete right hypoplastron; ETMNH-7640, right peripheral 10; ETMNH-7651, nearly complete left and right hyoplastra; ETMNH-7652, complete right costal 3; ETMNH-7670, left peripheral 10 and indeterminate limb bone fragment; ETMNH-7671, neural 2; ETMNH-7673, left costal 6; ETMNH-7677, complete left epiplastron; ETMNH-7678, right costal 1 and right peripherals 8–9; ETMNH-11744, left peripherals 9–10 and complete phalanx; ETMNH-12846, right peripheral 3; ETMNH-12978, portions or right costals 2–5; ETMNH-12989, complete right costal 3; ETMNH-12990, right peripheral 5; ETMNH-12992, right peripheral 7; ETMNH-13271, right peripheral 7; ETMNH-13281, left peripheral 5; ETMNH-13282, incomplete pygal; ETMNH-13283, left peripheral 10; ETMNH-14165, incomplete peripheral 10; ETMNH-14399, costal fragment; ETMNH-14556, nearly complete right costal 4; ETMNH-15948, nearly complete right hyoplastron; ETMNH-15949, right peripheral 8; ETMNH-17273, left peripheral 8; ETMNH-17274, costal fragment; ETMNH-17257, incomplete right costal 6; ETMNH-17349, right peripheral 9.
Type locality: Gray Fossil Site, Washington County, Tennessee, USA ( Fig. 1 View Figure 1 ) .
Type horizon and age: Latest Miocene-Early Pliocene (latest Hemphillian-Early Blancan NALMA). This estimate means the fossil locality, and C. corniculata , lies somewhere between Hh3 and Hh4, with recent age estimates based on biostratigraphy placing the site between 4.9–4.5 Mya.
Etymology: From the Latin corniculum, deminutive of cornu, horn, for example, with small horns, referring to the anterior projections of the carapace, specifically on the nuchal, and sometimes referred to as the nuchal horns. The common name of this species is: horned painted turtle.
Diagnosis: Chrysemys corniculata is placed in Emydidae due to the absence of musk ducts (would be notches on peripherals 3 and 4 if present), inframarginals reduced to two, normal hexagonal neurals 2–8 (also occurs in a few geoemydids; e.g. Mauremys Gray, 1869 ), and costal-inguinal buttress confined to costal 5. It is placed in the Deirochelyinae due to the lack of pectoral overlap of the entoplastron and lack of a hingeable plastral lobe with a ligamentous bridge connection (also present in some emydines). Diagnosed as a member of the genus Chrysemys on the basis of a low domed shell; anal notch absent or very weak; notching between posterior peripherals and marginals absent; dorsal carapacial keel absent; vertebral 1 with relatively slight nuchal overlap and with anterolateral flaring/projections. Distinguished from all other Chrysemys by extreme anterior projections of nuchal under marginals 1; anterior-most point of nuchal in middle of anterolateral projections (or anterolateral nuchal horns); concavity of the posterior edge of the cervical scute on the visceral surface; posterior inflation of vertebral 1; shorter and posteriorly shifted bridge with the axillary buttress barely contacting the posterior of peripheral 3 and the inguinal buttress contacting the posterior of peripheral 7; relatively shorter femoral scute along the sagittal midline; thick overlap of plastral scutes (i.e. where the scute wraps onto the visceral surface of the plastron), particularly of the humeral and femoral scutes; anterior projection of the epiplastron beneath the gular resulting in a mid-length indent along the anterior plastral lobe rim; strongly scooped, or ventrally dipping medial portion of the epiplastra; inflation of posterior plastral lobe under femoral scutes; indent at lateral plastral edge of femoral scute-anal scute sulcus contact; flattened posterior edge of xiphiplastra; shortened fossa orbitalis; anteroposteriorly shortened and curved cavum tympani; pronounced depression immediately dorsal to processus articularis; wider angled lower jaws (86° in C. corniculata vs. 74–78° in modern Chrysemys ); more pronounced sulcus cartilaginis meckelii; and more pronounced median ridge of the lower triturating surfaces.
Description
Methods: Terminology used throughout this study follows several well-known previous studies, including Thomson (1932), Zangerl (1969), Gaffney (1972), Ernst & Barbour (1989), Joyce & Bell (2004), Joyce (2007) and Jasinski (2018a). Although some previous studies have termed the keratinous, soft-tissue covering of the turtle shell as scales (e.g. Gaffney et al., 1998; Gilbert et al., 2001), we now know that the shell is covered with modified scales called scutes (e.g. Moustakas-Verho et al., 2014, 2017; Moustakas-Verho & Cherepanov, 2015) and therefore this latter term is used in the present study. Measurements are all maximum lengths and/ or widths unless otherwise stated. Orientations are in proper anatomical position unless otherwise stated. The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/.
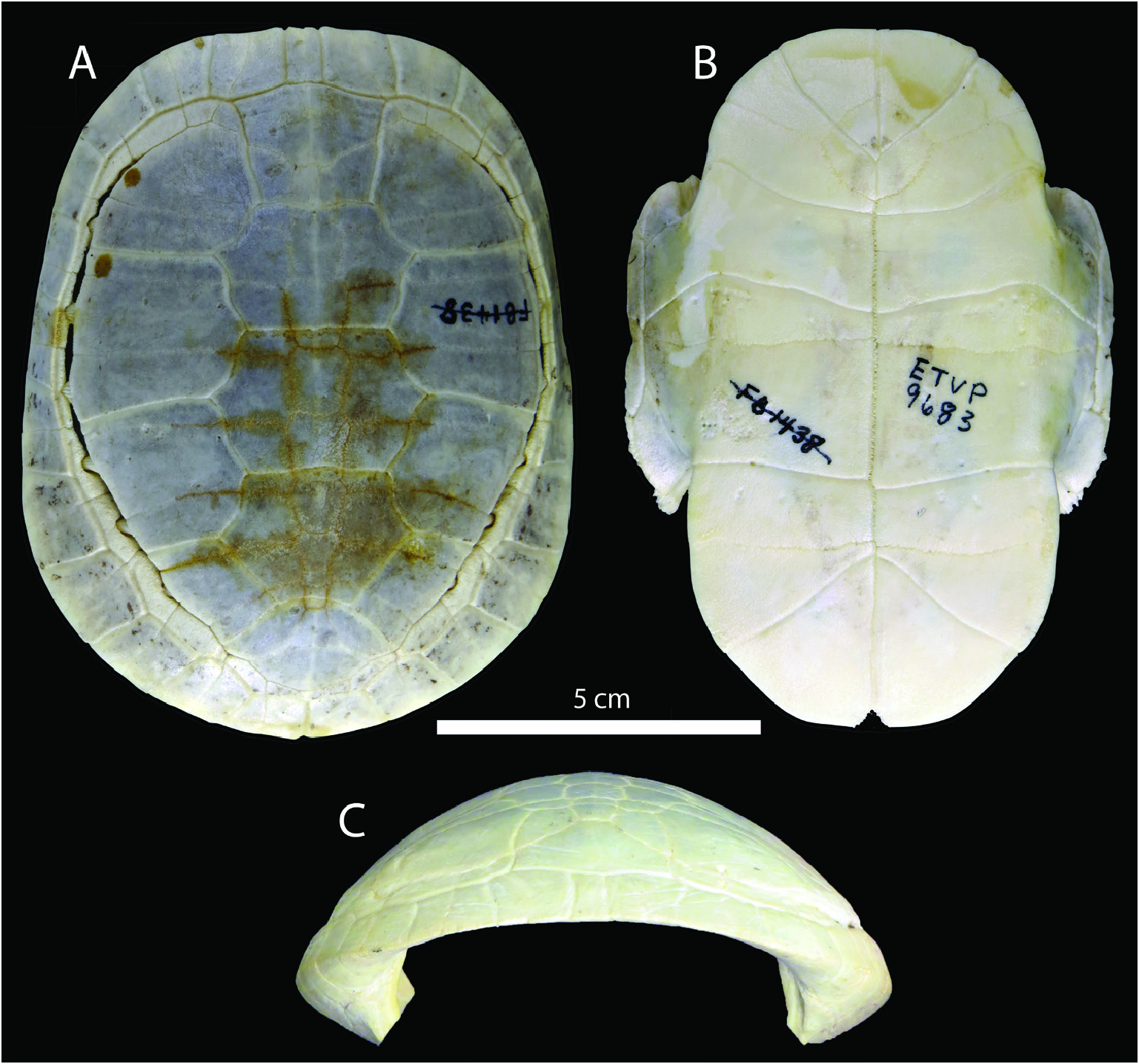
Shell ( Figs 2–3 View Figure 2 View Figure 3 , 5 View Figure 5 ): Chrysemys corniculata from the GFS is represented by multiple well preserved and mostly three-dimensional shells (including both carapaces and plastra). Although the specimens are sometimes somewhat crushed or ‘deformed’ while in situ, careful preparation often allows them to be re-assembled in their three-dimensional forms. The largest shells show maximum lengths (straight carapace length = SCL) of between 17.6 (ETMNH-12491) and 19.0 cm (ETMNH-20544), compared to C. picta , whose upper size (for females) is around 25.0 cm, although males achieve maximum size at around 15.3 cm ( Ernst & Lovich, 2009). The shell is more oval than in modern Chrysemys ( Fig. 4 View Figure 4 ), which can be more rounded, although some individuals can be more oval, particularly in older individuals of modern Chrysemys .
Carapace ( Figs 2–3 View Figure 2 View Figure 3 ): There are multiple wellpreserved carapaces, either partial or nearly complete. These give a good indication of the general size, shape and characteristics of C. corniculata . ETMNH-12491 is a medium-sized individual, but one that has reached adulthood based on fusion of the shell (particularly carapacial) elements (e.g. Legler, 1960; Zug, 1991), measuring 17.6 cm sagittally (SCL). ETMNH-20544 is the largest individual recovered, represented by a nearly complete shell with a few cranial and postcranial fragments, and measuring 19.03 cm (SCL). ETMNH-20609 represents the smallest complete (to nearly complete) specimen, with a carapace length of 14.03 cm (SCL). The shell has fine, inconspicuous texturing, although this cannot be physically felt and can normally only been seen when held to low angles with light ( Fig. 3 View Figure 3 C-D). This inconspicuous texturing is present in modern Chrysemys , albeit even more subtle ( Fig. 3 View Figure 3 E-F), and is not present in some other fossil Chrysemys taxa such as Chrysemys antiqua Clark, 1937 and Chrysemys timida Hay, 1908 . No median dorsal carapacial keel is present, and the posterior of the shell tends to have a sharp, downturned bend to it, making the posterior portion of the carapace nearly vertical ( Fig. 3B View Figure 3 ). This downturn can be present in modern Chrysemys , although it is always at a shallower angle than in C. corniculata . As noted above, the carapace of C. corniculata is more oval (regardless of size) than modern C. picta , which tend to exhibit a relatively rounded or circular carapace in dorsal view (length/width ratios of 1.40–1.48 in C. corniculata compared to 1.29–1.36 in C. picta ). In C. corniculata , the carapace also has a characteristic doming or raised portion at the anteromedial portion of the shell ( Fig. 3A View Figure 3 ), meaning the nuchal is noticeably higher than the surrounding elements, mainly the peripherals and corresponding marginals lateral to it. This is most easily seen in anterior view, where modern Chrysemys are usually gently rounded, while there is an open angle between the bridge and the sagittal anterior midline. Simply put, it is analogous to ‘circular vs. triangular’ in anterior view. Additionally, the entire bridge is shifted posteriorly in C. corniculata . This is evidenced by the axillary buttress barely contacting the posterior of peripheral 3 in C. corniculata ( Fig. 3B View Figure 3 ), while contacting the anterior of peripheral 3 in modern C. picta . For the inguinal buttress, both taxa contact the posterior of peripheral 7, indicating the bridge is a bit longer, larger and more robust in modern C. picta .
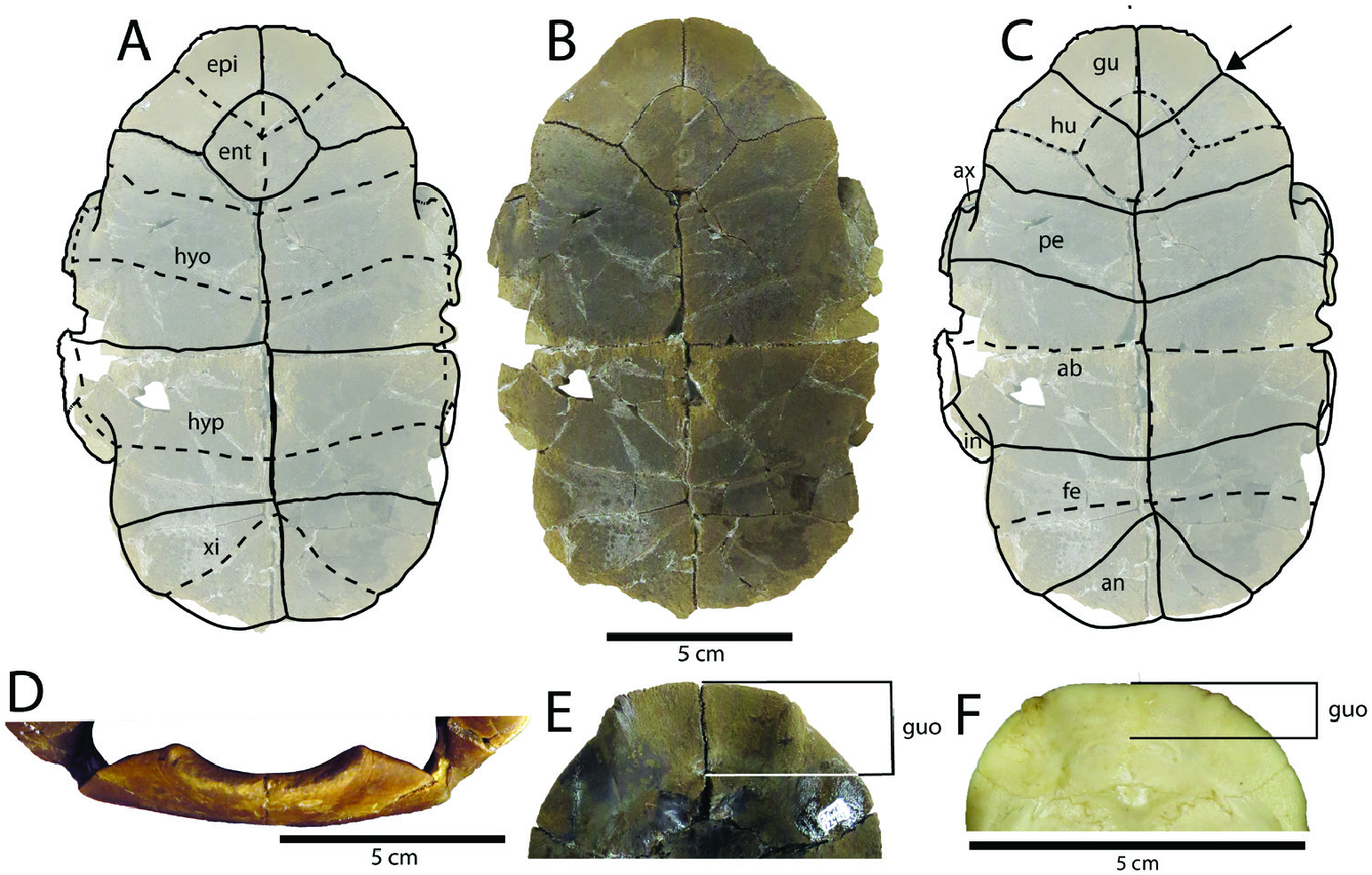
Sutures of the carapace ( Fig. 2A View Figure 2 ): The nuchal of C. corniculata represents a more pronounced and extreme morphology than that of modern Chrysemys . It maintains the roughly hexagonal shape of most emydid nuchals. The nuchal is nearly equal in length and width, although it is slightly longer than wide in ETMNH-12491 ( 42.4 mm long vs. 39.3 mm wide). Its anterior portion (anterior to the nuchal-peripheral 1-costal 1 intersection, 75% of nuchal length) is definitively longer than its posterior portion (25%). It is transversely constricted near the middle of the marginals 1, and flares anteriorly in C. corniculata , more so than in modern C. picta . The portion under the cervical scute does not reach as far anteriorly as that under the marginals 1, and these three projections are separated by two prevalent notches. The lateral edges of the anterolateral projections of the nuchal (nuchal horns) in C. corniculata are posterolaterally angled, making the anterior-most portion of the nuchal in the middle of these anterolateral projections, while there is no posterolateral-angling in other Chrysemys , including Chrysemys isoni Weems & George, 2013 . As stated above, the doming or raised region at the anteromedial portion of the shell means the nuchal is noticeably higher than the rest of the surrounding elements, mainly the marginals lateral to it ( Fig. 3A View Figure 3 ). This is most clearly seen in anterior view, where modern C. picta tend to be gently rounded ( Fig. 4C View Figure 4 ), while the anterolateral edges of the carapace are angled dorsally to the nuchal (from the bridge). The majority of the eight neurals agree morphologically with those in modern Chrysemys . Neural 2 is squat in C. corniculata with a significantly rounded posterior border, while the posterior border in modern C. picta is straight. Neural 8 is more squared-off than modern C. picta , which tend to have a more ‘triangular’ neural 8, although the neurals in turtles tend to have minor quantitative variations (e.g. Cherepanov, 1994). The posterior suprapygal, immediately anterior to the pygal, is concave on its anterior border, with a wider than long hexagonal shape. The anterior border of the posterior suprapygal in modern C. picta tends to be straight (mediolaterally or transversely oriented), but can be slightly concave, although not to the same extent as C. corniculata . While in modern C. picta the pygal routinely has parallel lateral edges, in C. corniculata it is contracted posteriorly, being narrower posteriorly than anteriorly. The pygal also possesses a small but conspicuous notch posteromedially with sharp edges. While an inconspicuous notch can be found in some modern Chrysemys , the notch in the latter has gently sloping sides and is relatively smaller. The anterior edge of the pygal exhibits a gentle and inconspicuous concave curvature. The bridge peripherals of C. corniculata are vertical, making them appear narrow when viewed dorsally. This mostly agrees with Recent Chrysemys , although those in the modern taxa are not as extreme, and therefore not as inconspicuous when viewed dorsally. The other peripherals agree morphologically with other Chrysemys . The edge of the carapace is smooth, with only inconspicuous notches between the posterior peripherals of some individuals (e.g. ETMNH-3561, -20609), not including those notches on and immediately around the nuchal. The outer rim is also relatively gracile, especially in comparison to the other deirochelyines known from the GFS ( Trachemys haugrudi ; Jasinski, 2018a). The axillary buttress barely contacts the posterior of marginal 3 in C. corniculata ( Fig. 3B View Figure 3 ), while it contacts the anterior of peripheral 3 in modern C. picta . The inguinal buttresses of both taxa usually contact the posterior of peripheral 7. This suggests a relatively shorter and potentially less robust bridge for C. corniculata . As noted above, the bridge peripherals are oriented mainly dorsoventrally, making them more visible laterally and less significant when viewed dorsally or ventrally. This leads to the posterior portion of the carapace flaring out laterally beginning midway through peripheral 7, as in modern Chrysemys .
Sulci of the carapace ( Fig. 2C View Figure 2 ): The bony shell of a turtle is covered by keratinous scales or plates called scutes. Scutes are modified scales and develop differently than reptilian scales, namely with the former developing from local epithelial thickenings called placodes through radial growth while scales (along with feathers, teeth and hair) develop as ectodermal appendages through epithelial-mesenchymal interaction (e.g. Cherepanov, 2006; Moustakas-Verho et al., 2014, 2017; Moustakas-Verho & Cherepanov, 2015). These scutes are separated into various different sections by sulci (or seams), which can be seen on the surface of the bones and give an indication of their morphology and appearance even when they are not present or preserved, such as in fossils. The cervical scute is long and narrow, often coming to an anterior point sagittally (medioanteriorly). The cervical underlap is shorter than the overlap in C. corniculata (~74–85%), although less so than in modern C. picta (~67–80%), but these values tend to vary, as seen by the range. The posterior width of the overlap of the cervical scute is narrower (compared to its length) in C. corniculata than either C. isoni or C. picta . The lateral edges of the cervical are parallel to slightly angled anteriorly as the cervical comes to an anterior point. As stated above, it is separated from marginals 1 by a pronounced lateral notch. The anterior-most point(s) of the carapace lie medially on the anterior projecting portion of marginals 1 (anterolateral projections of the nuchal under marginals 1). Although vertebrals 2–5 agree morphologically with those in C. picta , vertebral 1 is more morphologically unique ( Fig. 3C View Figure 3 ). In C. corniculata , vertebral 1 is straight anteriorly and well-rounded (concave) posteriorly. There is lateral inflation posteriorly while it is constricted anteriorly. At its anterolateral edges vertebral 1 flares laterally and becomes significantly wider, giving it a slightly hourglass-like shape. However, in modern C. picta , the anterior and posterior borders of vertebral 1 tend to be relatively straight to slightly curved, while the anterior half is normally wider (the sides can also sometimes be approximately parallel) ( Fig. 3E View Figure 3 ). Regardless, the only unique curvature present is at the anterolateral points, which curve slightly laterally. Vertebral 4 also exhibits more curvature and has more sinusoidal sulci, although this is mainly visible posterolaterally where it contacts pleural 4. As in other Chrysemys , and emydids in general, C. corniculata has four pairs of pleurals that mostly agree morphologically with those of modern Chrysemys . However, as for the morphology of the vertebrals, the medial sulci of pleurals 1 and 4 in C. corniculata are more sinuous and curved where they contact vertebrals 1 and 4, respectively. As noted above, marginal 1 has a pronounced medial projection, clearly distinguishing its anteromedial and anterolateral halves ( Figs 2 View Figure 2 , 7 View Figure 7 A-D). The anterior extent of this projection tends to be pronounced, although it is less so in ETMNH-12491. This difference may be individual variation or sexually dimorphic, although it is present to at least some degree in all nuchal specimens of C. corniculata . There is a small, inconspicuous indent on marginal 4 at the anterior edge of the bridge. The carapace flares out laterally and become wider at the sulci between marginals 7 and 8, making this the widest part of the carapace. The rest of the marginals agree morphologically with those in C. picta , although there can be inconspicuous notches at the rim of the posterior marginals of C. corniculata . The posterior-most marginals (=supracaudal scutes of some authors) possess a small, but noticeable, notch posteromedially and do not reach onto the last suprapygal.
Plastron ( Fig. 5 View Figure 5 ): The plastron follows the general emydid shape and composition with two epiplastra, an entoplastron, two hyoplastra, two hypoplastra and two xiphiplastra. In ETMNH-20544, the complete plastron measures approximately 17.25 cm anteroposteriorly (straight plastron length, SPL). The smallest nearly complete shell specimen (ETMNH-20609), has a sagittal plastron length of 12.54 cm. A suture runs medially through the plastron separating the two sides and the elements of the plastron that are paired. However, it does not run through the entoplastron, as is the case with other emydids and turtles in general. Both plastral lobes are inflated (laterally), although the posterior plastral lobe is significantly more so. The anterior plastral lobe bends dorsally (viscerally) toward its anterior extremity ( Fig. 3B View Figure 3 ).
Sutures of the plastron ( Fig. 5A View Figure 5 ): The epiplastron morphology agrees strongly with that in modern Chrysemys ( C. picta and its subspecies, including C. picta dorsalis ). The epiplastra exhibit a gentle curvature making up the anterior portion of the anterior plastral lobe. They exhibit an anterior projection medial to the gular-humeral sulci, which is more conspicuous than in modern Chrysemys . The entoplastron agrees in general morphology with modern Chrysemys and other deirochelyines. However, individual morphology tends to be variable, with some relatively uniform with the anterior and posterior halves approximately equal in length (in relation to the point where the lateral projections lie; e.g. ETMNH-3561, -20544 and -20609), while others are longer than wide (ETMNH-12491) or wider than long (ETMNH-3559). The morphology of the entoplastron can also be variable in modern C. picta , showing similar shape and dimensional variation in the species. The posterior border of the entoplastron is gently rounded, while in other Chrysemys it tends to be straight (mediolateral or transverse) or angled. The suture between the epi- and hyoplastra nearly bisects the anterior lobe, with no indent or notch present at its lateral edge. The hyo-hypoplastral contact is nearly straight, running transversely and bisecting the bridge. The hypo-xiphiplastral sutural contacts exhibit a gentle convex (anterior) curvature, with the lateral edges curving posteriorly (caudally). Although this contact is usually relatively straight in modern Chrysemys , it can exhibit some curvature in some C. picta individuals (e.g. ETVP 9672, Chrysemys picta bellii Gray, 1830 ).
Sulci of the plastron ( Fig. 5C View Figure 5 ): The surface of the plastron is covered with a number of scutes (or scales), of which the sulci (or seams) left behind can give an indication of their morphology and appearance. Chrysemys corniculata has the same complement of scutes as other Chrysemys with a pair of gular scutes, a pair of humeral scutes, a pair of pectoral scutes, a pair of abdominal scutes, a pair of femoral scutes and a pair of anal scutes. Anteroposterior lengths are measured sagittally for determining the plastral formulas. The overall plastral formula for C. corniculata is abdominal> anal> gular> pectoral> humeral> femoral. This is unique from modern C. picta , which is abdominal> anal> gular> pectoral> femoral> humeral. This is due to a relatively shorter femoral scute along the midline, rather than a relatively longer humeral, in C. corniculata .
The gular scutes reach approximately midway through the entoplastron, similar to those in modern Chrysemys . However, they extend farther anteriorly, creating a pronounced anterior projection beneath them. The anterior projection is confined to the epiplastron beneath the gulars. While C. picta has a rounded anterior plastral lobe with a continuous curve, the anterior projection of C. corniculata results in a discontinuous curvature, with the gulars projecting anteriorly and a concavity on the outer rim at the contact between the gulars and humerals ( Fig. 5C View Figure 5 ). In addition, in C. corniculata , the gular scute overlap is larger (10.1 to 12.6%) than in modern C. picta (6.9 to 7.4%) ( Fig. 5 View Figure 5 E-F). The lateral gular anterior projections (or gular horns) are separated from the rest of the anterior edge of the plastron by a small notch. As mentioned above, the gulars project dorsally (or viscerally, when viewed anteriorly), creating an obvious embayment ( Fig. 5D View Figure 5 ). The humeralpectoral scute sulci are straight to slightly concave (posteriorly), with their lateral-most edges curving more sharply posteriorly, similar to those in modern Chrysemys . The axillary scutes are relatively small and found on the anterior edge of the bridge, similar to their morphology and position in C. picta . The inguinal scutes, on the other hand, are enlarged and possess a sigmoidal curvature dorsally ( Fig. 3B View Figure 3 ). There is also a conspicuous posteroventral curvature between the inguinals and abdominals. This curvature is more pronounced than in modern Chrysemys and is present definitively on the bridge, while in modern Chrysemys it lies on the anterior-most portion of the posterior plastral lobe, making it more vertical in C. corniculata , vs. more horizontal (or lateral) in modern Chrysemys . The pectoral-abdominal sulci are concave posteriorly, more so than in C. picta . Similarly, the lateral edges also curve posteriorly as they near the bridge. The abdominal-femoral sulci have a gentle posterior concave curvature and a unique posterior curvature laterally as they near the posterior edge of the bridge. As in modern Chrysemys , the femoral-anal sulci are strongly convex posteriorly, with midline contact just posterior to the hypo-xiphiplastral sagittal midline contact and terminate on the posterolateral rim of the posterior plastral lobe. While the lateral edge between the femoral-anal sulci contact is relatively inconspicuous in modern Chrysemys , there is a unique and pronounced notch in C. corniculata . Posteriorly, as for the bones of the plastron, the posterior edge of the anals is straight in C. corniculata with no conspicuous notch noted at the midline contact ( Fig. 5 View Figure 5 A-C), while modern Chrysemys tends to possess a small but conspicuous notch ( Fig. 4B View Figure 4 ).
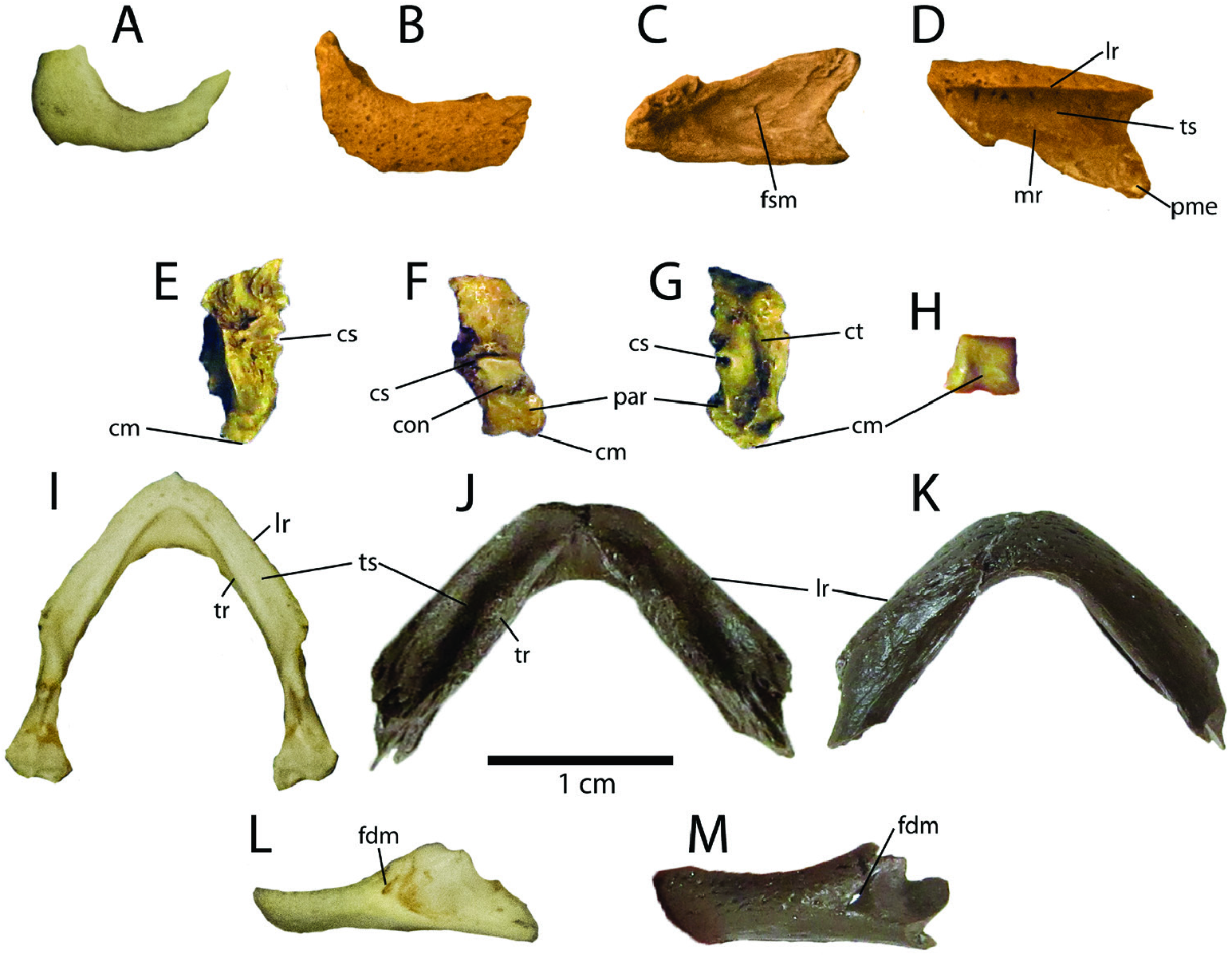
Skull ( Fig. 6 View Figure 6 ): Several cranial fragments of C. corniculata are currently known. These include a nearly complete left maxilla (ETMNH-20544), left and right quadrates (ETMNH-3559) and a nearly complete set of lower jaws (ETMNH-12491). None of these specimens are isolated and all include partial shells to ensure their referral to C. corniculata . While only a few cranial fragments are preserved, their preservation shows unique morphological features contrasting with modern Chrysemys . The left maxilla (ETMNH-20544) is missing its anterior portion that would articulate with the premaxilla ( Fig. 6 View Figure 6 B-D). However, it is still relatively short at 10.8 mm [maximum length of the external (lateral) surface]. A small portion of the caudolateral surface is also broken. This bone preserves a ventral portion of the fossa orbitalis, and the sutural surface for the jugal has a ‘v’-shaped or triangular attachment surface. This attachment is similar to that in modern Chrysemys , but the sutural surface (and therefore the jugal) are more rostral and ventral in C. corniculata , leading to a smaller fossa orbitalis and orbit. Similar to C. picta , the foramen supramaxillare lies at approximately the same level as the anterior extent of the maxilla-jugal sutural contact dorsally ( Fig. 6C View Figure 6 ), but in C. corniculata it is enlarged and more conspicuous. The posteromedial extension of the maxilla is robust and rounded in C. corniculata , although it would appear to have a stronger apex if it was attached to the jugal. Ventrally (viscerally), there are several foramina along the triturating surface medial to the labial ridge. These foramina sit between a raised median ridge that runs through the middle of the triturating surface and the labial ridge. These foramina lie in a shallow depression, as in modern Chrysemys , although this depression becomes more pronounced posteriorly in C. corniculata compared to modern Chrysemys . The triturating surface of C. corniculata is also relatively wider in C. corniculata than in C. picta , while the median ridge is more pronounced in the former as well.
The quadrates are isolated but well preserved in ETMNH-3559 ( Fig. 6 View Figure 6 E-H). Both the left and right quadrates are nearly complete and preserve the condylus mandibularis (or mandibular condyle) and a significant amount of the cavum tympani. The left quadrate has a maximum dorsoventral preserved height of 9.9 mm, while the right is 11.2 mm. Caudoventrally the processus articularis has a similar trapezoidal morphology in both C. corniculata and C. picta . The condylus mandibularis is similar in morphology to that of modern C. picta , although its concavity, located anteroventrally on the processus articularis, is less conspicuous. The processus articularis has dimensions of 3.8 × 3.7 mm. A small portion of the canalis system is preserved with both quadrates, although part has been sheared off and is incomplete, leading to difficulty determining which portions of the mainly pneumatic canalis systems are preserved. There is a deep, pronounced concavity immediately rostral (anterior) to the condylus mandibularis that is more pronounced in C. corniculata than in adult specimens of modern C. picta (e.g. ETVP 9672). However, this concavity appears to be more pronounced in ontogenetically younger C. picta specimens (e.g. ETVP 9691), implying the concavity is ontogenetically variable. Regardless, this concavity is deeper in C. corniculata than in even ontogenetically young modern C. picta , potentially suggesting a deeper concavity is ancestral in Chrysemys . A foramen is present dorsal to this concavity on the rostral surface of the quadrate. This foramen is also present in modern Chrysemys , although it is situated more lateral in C. corniculata . The morphology of the cavum tympani can also be partially derived from the quadrates. While in modern Chrysemys the cavum tympani is well rounded, in C. corniculata it would have been rostrocaudally compressed, making it more oval. A slight curve toward the middle portion makes it appear as a crescent. A rostrodorsal compression of the cavum tympani is also seen in ontogenetically younger modern Chrysemys specimens, also suggesting a rostrodorsally compressed cavum tympani is ancestral in Chrysemys . The cavum tympani appear to anteroposteriorly inflate and become more rounded as individuals age. However, this curvature is not present in modern Chrysemys . Even if ETMNH-3559 is an ontogenetically younger individual and some of the morphology of the quadrates would change in older individuals, there are still morphological distinctions regardless of the ontogenetic ages or stages being compared.
A nearly complete set of lower jaws is preserved with the holotype ( ETMNH-12491, Fig. 6 View Figure 6 J-K, M). The lower jaws consist of the right and left dentaries, although most of the caudal elements of the lower jaws were not recovered. The jaws have a maximum rostrocaudal length of 12.1 mm, a maximum overall length of 16.1 mm (measured from apex along right dentary to its distal point), and a maximum preserved width of 19.9 mm caudally. The angle of the lower jaws (in dorsal and ventral views) is larger in C. corniculata compared to modern Chrysemys . Indeed, the angle of the lower jaws is 86° in the former, but only approximately 74–78° in modern C. picta . These wider jaws suggest a wider skull in C. corniculata as well. Rostrally (anteriorly), the hook on the rostromedial portion of the labial ridge is not preserved. A tomial (or medial) ridge, more pronounced than in modern Chrysemys , is present on the lower triturating surface, with a shallow depression between the tomial and labial ridges. The labial ridge is smooth as in other Chrysemys . The lower triturating surface is relatively wider in C. corniculata than in modern Chrysemys , but not significantly so. The sulcus cartilaginis meckelii is pronounced, particularly since the tomial ridge provides a conspicuous ledge for it dorsally. While it is more prominent than in modern Chrysemys , it is significantly less than that in Pseudemys . The foramen alveolare inferius is pronounced and is larger than in modern C. picta . Laterally, the foramen dentofaciale majus is more pronounced in C. corniculata compared to C. picta , although the lateral overhang from the caudal portion of the labial ridge is less pronounced in the former.
Postcranial skeleton: Aside from the shell, postcrania are present in several specimens, although almost all material is fragmentary and incomplete. This is probably due to the palaeoenvironment and taphonomic conditions as other animals and organisms would have had opportunities to disarticulate and/ or break bones before burial. Additionally, it is common to have non-shell material washed away from the shells in aquatic environments (e.g. Brand et al., 2003). Specimens with at least some postcrania include ETMNH-3559, ETMNH-3561, ETMNH-7670, ETMNH-11744, ETMNH-12491 and ETMNH-20544.
Axial skeleton: A single cervical vertebra is preserved with ETMNH-20544, identified as an incomplete cervical vertebra 7. It has a preserved maximum axial length of 7.8 mm through the centrum. The cranial centrum surface has two concave surfaces transverse to each other, as in other emydids. The prezygapophyses are not preserved. Dorsally, the neural spine is split. There is a point cranially where the two halves meet, located just caudal to where the prezygapophyses would lie. The two halves project caudally onto the two prominent postzygapophyses. They form a conspicuous ‘V’-shaped ridge that points cranially. There is also a significant depression within the ‘V’ and craniomedial to the postzygapophyses. The articular surface of the postzygapophyses lies flat (craniolaterally) and points ventrally. Ventrally, the hypapophysis is prominent, although it is broken cranially. The hypapophysis is also prominent in modern Chrysemys , although its development varies throughout the cervical vertebral series. The centrum is constricted at its mid-length. Cranioventrally there are two prominent ‘knobs’ beneath the two anterior concavities of the cranial centrum surface. These ‘knobs’ are more well-defined than those in modern Chrysemys . There is no trace of the caudal ‘knobs’ and resultant ridges present in the more cranial (or anterior) cervical vertebrae and, specifically, in cervical VI in modern Chrysemys . The neural canal is subtriangular. Several faint foramina are present, although the foramina located just caudal to the prezygapophyses are by far the most prominent. ETMNH-12832 has a cervical vertebra VII that is 13.4 mm long (axially) and agrees morphologically with that of ETMNH-8549. Dorsal vertebral fragments are preserved still attached to the carapaces of some specimens. These are triangular with constricted bases (ventrally) and agree morphologically with other Chrysemys specimens. There are no caudal vertebrae preserved with any of the specimens yet recovered.
Appendicular skeleton
Forelimbs: A small proximal fragment of a left coracoid of C. corniculata is preserved with ETMNH-12491. The maximum width is 9.1 mm across the articular surfaces near the glenoid fossa. The proximal end has three articular surfaces. The articulation with the scapula and acromial process is a ‘half-circle’ with an inflated ridge running through its middle, while the portion of the glenoid fossa from the coracoid is sub-triangular. This morphology agrees with that in modern Chrysemys . A relatively well preserved, but incomplete, right humerus is preserved with ETMNH-20544. The proximal and distal portions are preserved, while the middle portion of the shaft is not. Proximally, the medial tuberosity is significantly larger and more prominent than the lateral tuberosity. The humeral head is oval, similar to modern Chrysemys . There is a marked ridge lateral (external) to the humeral head. While this protuberance is also present in modern Chrysemys , it is less conspicuous, with the ridge creating an obvious groove between the lateral protuberance and the humeral head in C. corniculata . Distally the humerus in C. corniculata has similar morphology to that in modern Chrysemys . There is a recognizable groove for the ulnar nerve leading to a small but conspicuous foramen on the lateral surface near the distal end. While this feature is also seen in modern Chrysemys , the groove leading to the foramen is more prominent in C. corniculata . Humeral fragments preserved with ETMNH-3561 and -12491, along with a highly fragmentary left humerus with ETMNH-20544, agree with those features listed above. A complete right radius is preserved with ETMNH-3559. It has a maximum length of 13.53 mm, with diameters of 2.37 mm at its proximal end, 3.47 mm at its distal end and 1.34 mm at mid shaft. The proximal surface is flattened and semi-circular. Its medial edge is flattened at the proximal surface, and there is a depression on the medial surface near the proximal end. The distal end is twisted compared to the proximal end, with the ‘oval’ shape significantly wider transversely at the proximal end. The shaft flares out near both the proximal and distal ends. The lateral edge projects farther distally than the medial edge. There is also a slight depression on the shaft near the distal end on both the dorsal and ventral surfaces which, while also present in modern Chrysemys , is inconspicuous in the latter, while more prominent in C. corniculata . Two indeterminate carpals are preserved with ETMNH-3559. They are both small elements that are rounded and relatively flattened proximodistally. Little else can be said of them or their morphology. Three manual phalanges are also preserved with ETMNH-3559, which adhere to the general phalangeal morphology of emydids and most other turtles. ETMNH-3559 also preserves three manual unguals, although only one is complete. This ungual is small, with a length of 5.3 mm and a proximal diameter of 1.6 mm. It possesses lateral grooves, which are relatively prominent, particularly proximally, with some tuberosity around the edges of the proximal surface. A single ungual was collected with ETMNH-20544, although it is uncertain whether it represents a manual or pedal ungual. The small claw has a length of 6.3 mm and a proximal diameter of 1.9 mm. There is also some tuberosity around the edges of the proximal surface. While little work has been done with emydid claws, illustrations provided by Jasinski (2018a) of a fossil emydid ( Trachemys haugrudi ) hinted that the lateral grooves are more pronounced in manual unguals. The lateral grooves on the current ungual are small, but deep and pronounced, suggesting this represents a manual ungual. Other forelimb material, including the scapula, ulna and metacarpals, are currently unknown, although they are presumed to be similar to those elements in modern Chrysemys .
Hindlimbs: Small pelvic fragments of C. corniculata are present in ETMNH-20544. The proximal portion of a right ischium is preserved. The proximal end has a maximum length of 9.48 cm across the articular surfaces. Theboneisgracile, particularlyjustdistaltothearticular surfaces and strongly agrees with the morphology in modern Chrysemys . A small fragment of a left femur is preserved with ETMNH-3559. It represents part of the proximal end and, while incomplete, does provide some clues as to its overall morphology. Both tuberosities on the proximal end would have been relatively small, similar to those in modern Chrysemys . A portion of the femoral head is preserved and suggests it would be largely offset. It would have extended significantly from the femoral shaft, probably more so than in modern Chrysemys . Additionally, while incomplete, the femoral head is inferred to have been relatively small as well. A complete right fibula is preserved with ETMNH-12491 with a maximum proximodistal length of 22.1 mm. It is a thin, gracile element similar to that of modern Chrysemys , with widths of 1.8 mm at its proximal end and 4.1 mm at its distal end. Proximally, the element is nearly round, thin and inconspicuous, before it begins to flare out towards the distal end. Distally, the surface is sub-oval. There is a subdued ridge that runs laterally along its length before reaching the distal surface, where it becomes more pronounced. A single, isolated phalanx is preserved with ETMNH-11744. While it is not known whether this comes from the manus or pes, it generally agrees morphologically with phalanges of other Chrysemys , and other emydids. As mentioned above, while an ungual is known, it is currently inferred to be a manual ungual due to prominent lateral grooves. Other pelvic and hindlimb material, including the ilium, pubis, tibia, metatarsals and tarsals, are currently unknown.
FURTHER COMPARISONS
Trachemys antiqua [ holotype = YPM (PU) 13839] is based on a nearly complete plastron and carapace fragments from the Middle Chadron Formation (Middle Chadronian NALMA) in Pennington County, South Dakota, USA ( Clark, 1937). It was re-evaluated by Hutchison (1996), who referred it to Chrysemys . However, Joyce et al. (2013) questioned its placement in Chrysemys , and Vlachos (2018) agreed with this more recent assessment. While Joyce et al. (2013) identified the species as in ‘ Chrysemys ’, Vlachos (2018) identified it as in ‘ Trachemys ’. Regardless of which, if any, of these ambivalent referrals is correct, it will remain in Chrysemys in the present study until further information is known about its phylogenetic and generic placement. Chrysemys corniculata differs from C. antiqua in several ways, most dealing with the plastron as the carapacial fragments provide few useful characteristics. The gular region of the anterior plastron of C. corniculata projects anteriorly, particularly at the gular-humeral sulcus, while this region in C. antiqua maintains a gentle curvature of the anterior plastral lobe. The lateral edges of the pectoral-abdominal sulci are not as strongly curved as in C. antiqua . The abdominal-femoral sulci are concave, while in C. antiqua they are straight to roughly flat. The entoplastron is well-rounded, particularly posteriorly, in C. corniculata vs. straighter to angled in C. antiqua . While Chrysemys exhibit a reduced anal notch in the posterior plastral lobe, it is even further reduced in C. corniculata than in other species, including in relation to C. antiqua . While the bridge peripherals in C. corniculata are laterally thin, and oriented mainly dorsoventrally, those of C. antiqua flare laterally more, making them more visible when viewed dorsally (and ventrally) in the latter. The posterior carapace of C. corniculata also flares laterally at the level of peripheral 7, whereas C. antiqua does not exhibit posterior flaring.
Chrysemys isoni ( holotype = CMM-V-4664) was identified from an incomplete nuchal from the Upper Fairhaven Member (Fairhaven Bed C, Late Early Hemingfordian) of the Calvert Formation at the boundary of King William and New Kent counties, Virginia, USA ( Weems & George, 2013). The type nuchal was referred to Chrysemys based on several features, including aspects of its anterior and lateral borders, and the cervical scute ( Weems & George, 2013). While Vlachos (2018) considered this species a nomen dubium, features of the nuchal are still unique, allowing for a diagnosis of the species. A key feature of C. isoni is an elongate underlap of its cervical scute in relation to its overlap, suggesting a longer cervical scute underlap than other known deirochelyines. This relationship in C. corniculata is more similar to modern C. picta and Deirochelys reticularia Latreille in Sonnini & Latrielle, 1801 than to C. isoni . Additionally, the posterior width of the overlap of the cervical scute is shorter (compared to its length) in C. corniculata than either C. isoni or C. picta . The lateral edges of the anterolateral projections of the nuchal that would lie under marginals 1 in C. corniculata are angled posterolaterally, making the anterior-most portion of the nuchal in the middle of these anterolateral projections or horns, while there is no posterolateral-angling in C. isoni , making the anterior-most point at the lateral edges of these anterolateral projections in the latter. This latter characteristic is also seen in modern C. picta and other deirochelyines. Finally, the nuchal is more gracile in C. corniculata but definitively more robust in C. isoni .
Chrysemys timida [ holotype = YPM (PU) 10853] is based on an incomplete carapace and plastron from the ‘ Equus beds of … Nebraska’ ( Hay, 1908: p. 345) making it Pleistocene in age. Vlachos (2018) recently determined the species was valid, but its placement within Chrysemys was less certain, and identified is as ‘ Chrysemys ’ timida . Until further details on its phylogenetic or generic placement are determined, the species is retained in Chrysemys here. Hutchison (1996) determined C. timida and C. antiqua were similar, with only a few differences from the overlapping material. One of these features was the presence of three suprapygals in C. timida , rather than two in C. antiqua , and C. corniculata agrees with the latter in this regard. The posterior-most suprapygal is also distinct in the two taxa, being sub-hexagonal in C. corniculata with a concave anterior border, and while it is also sub-hexagonal in C. timida , the anterior border is markedly convex in the latter. The neural series bones are more squat in C. corniculata , tending to be shorter, whereas those in C. timida are elongate. The point of contact between vertebral 1-pleural 1-marginal 1 lies on peripherals 1 in C. timida , the nuchal in C. antiqua , and on the nuchal just medial to the nuchalperipheral 1 suture in C. corniculata . The nuchal of C. timida is also almost equally divided (lengthwise) between the lateral points of contact of nuchal-peripheral 1-costal 1, whereas in C. corniculata the anterior portion is noticeably longer than the posterior portion. The type specimen of C. timida is in poor condition, making determining other characteristics difficult.
The anterior carapacial morphology of C. corniculata is distinct from other Chrysemys ( Fig. 7 View Figure 7 ). However, anterior projections of the shell, particularly of marginals 1, have been mentioned in modern C. picta , particularly in a population of Chrysemys picta marginata Agassiz, 1857 by Moldowan et al. (2020a, b). While these projections are present exterior to where the nuchal horns would be, those in modern Chrysemys are soft-tissue features present on the scutes ( Fig. 7G View Figure 7 ) and are far less conspicuous on the nuchal ( Fig. 7 View Figure 7 E-F). The nuchal horns are present on all specimens of C. corniculata , although some are larger and project farther ( Fig. 7 View Figure 7 C-D) than others ( Fig. 7 View Figure 7 A-B). Therefore, this feature may be sexually dimorphic with larger nuchal horns present in males. The more extreme anterior projections of marginals 1 were noted in the males of C. p. marginata by Moldowan et al. (2020a, b). However, this feature is not pronounced osteologically ( Figs 4 View Figure 4 , 7 View Figure 7 E-G). This ‘weaponized anterior shell’, as termed by Moldowan et al. (2020b), specifically refers to the scutes and soft tissue of the shell and is an extreme exaggeration of the underlying osteology. This would have probably been present in C. corniculata , but to a more extreme degree as the marginals 1 region of the nuchal is more pronounced than in modern Chrysemys . Regardless, osteologically C. corniculata can be differentiated from other Chrysemys based on this region, among other features.
Two other species previously referred to Chrysemys likely do not belong to the genus. Chrysemys aeilliamsi ( holotype = UF 11561) is based on a nearly complete carapace and plastron from the Early Hemphillian McGehee Farm locality in Alachua County, Florida, USA ( Rose & Weaver, 1966). Although it was named Chrysemys aeilliamsi , it was later referred to Pseudemys (considered a subgenus at the time), making the original referral Chrysemys (subgenus Pseudemys ) aeilliamsi by Auffenberg (1972) (see also Hulbert, 2001). This referral was upheld by Vlachos (2018). The generic placement of this species was further called into question by Jasinski (2018a), and the current study finds its placement in Chrysemys (or Pseudemys ) improbable (see below).
Chrysemys carri ( holotype = UF 9427) was named based on a nearly complete carapace and plastron from the Early Hemphillian McGehee Farm locality in Alachua County, Florida, USA ( Rose & Weaver, 1966). Like ‘ C.’ aeilliamsi above, it was also referred to Pseudemys (considered a subgenus at the time) by Auffenberg (1972) (see also Hulbert, 2001), and this generic placement was upheld by Vlachos (2018). Jackson (1976) reviewed C. carri and determined it was synonymous with Pseudemys caelata Hay, 1908 . This allowed him to refer the latter species to Chrysemys as Chrysemys caelata , with C. carri as a junior synonym. Jasinski (2018a) suggested P. caelata was probably referable to Deirochelys , which implies both would be representative of Deirochelys . The current study finds a similar phylogenetic position, suggesting a similar generic placement as potentially Deirochelys . Further complicating matters is the fact that a fossil species of turtle was named Deirochelys carri Jackson, 1978 , although its generic placement has been recently questioned by Vlachos (2018). Therefore, a potential referral of ‘ Chrysemys ’ carri to Deirochelys would further complicate matters. A reassessment of the generic placement of both ‘ Chrysemys ’ carri and ‘ Pseudemys ’ caelata , along with that of ‘ Deirochelys ’ carri , will be undertaken in a future review of fossil emydids and their evolutionary relationships.
PHYLOGENETIC ANALYSIS
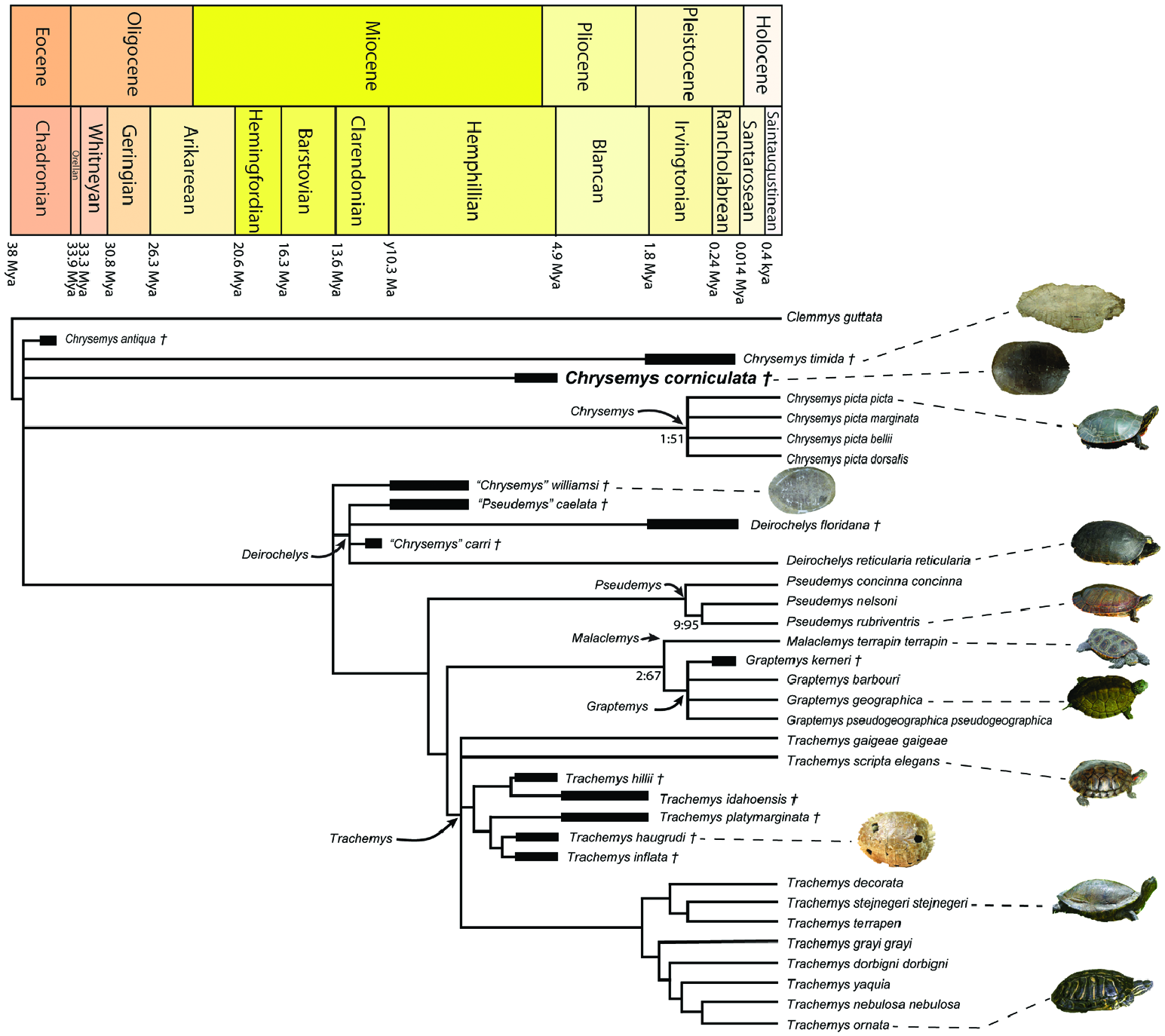
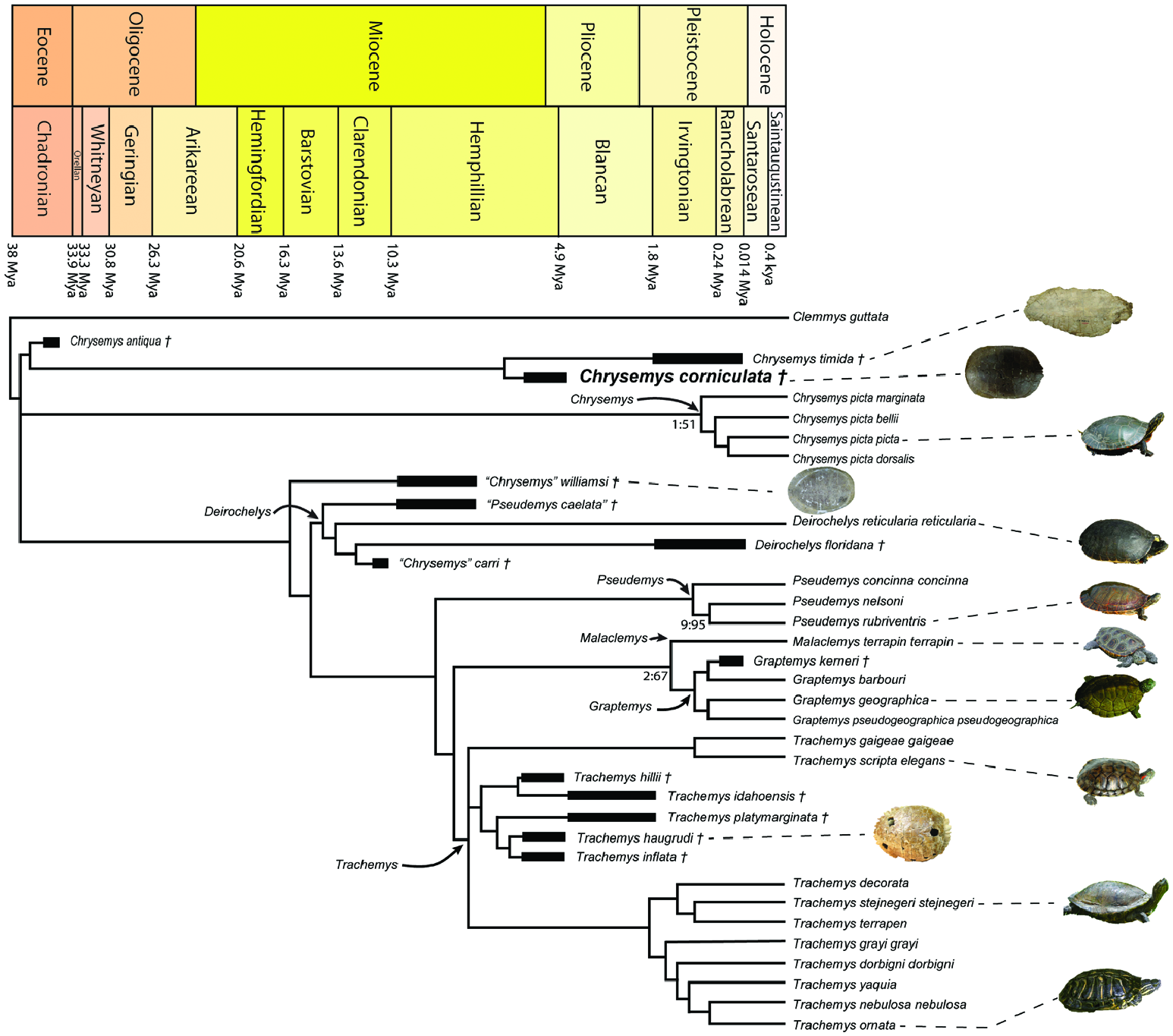
Chrysemys corniculata was integrated into a list of currently accepted extant deirochelyine taxa, along with several other suspected deirochelyine fossil representatives, to investigate its phylogenetic relationships. The analysis was based on the morphological character set of Jasinski (2018a), which was in turn was based on characters from numerous other earlier character sets and sources [see Jasinski (2018a) and references therein]. Additionally, three new characters were added to the dataset to help reflect some of the intergeneric morphological variation among Chrysemys . These characters (characters 244–246) focus on variation in plastral formula, and measurements for these are taken along the sagittal midline along the sagittal sulci (or seams). Juveniles were excluded from the dataset to avoid ontogenetic variation of characters. Variation is an important aspect to be taken into account when investigating biodiversity and taxonomy in fossil organisms (e.g. Lucas et al., 2011, 2016; Ponssa et al., 2011; Sullivan et al., 2011a, b, c, d, e; Sullivan & Jasinski, 2012; Cavin et al., 2013; Jasinski, 2013b, 2015a, b; McGarrity et al., 2013; Jasinski & Wallace, 2014, 2015; Dalman et al., 2017, 2021, 2022; Jasinski et al., 2020, 2022; Gee & Jasinski, 2021), including morphological changes through ontogeny (e.g. Jones & German, 2005; Jasinski, 2011; Jasinski & Sullivan, 2011, 2016; Sullivan et al., 2011c; Piro & Hospitaleche, 2019; Vamberger et al., 2020; Lichtig et al., 2021; Ishikawa et al., 2022) or sexual dimorphism (e.g. Litzgus & Smith, 2010; Vega & Stayton, 2011; Moldowan et al., 2016a, b). Although the amount of morphological variation in modern species can be quite high (e.g. Kennedy et al., 1980, 1986; de Schepper et al., 2004; Arribas et al., 2013; Watson et al., 2019; Brochu & Sumrall, 2020), this was taken into account by Jasinski (2018a) in the dataset by considering degrees of polymorphism in the character set. This was also accounted for with the scoring of the new operational taxonomic units (OTUs) in the current dataset, including C. corniculata .
Utilizing the larger dataset developed by Jasinski (2018a) allows for the interrelationships of Chrysemys , in addition to those specifically of C. corniculata , to be investigated. To better focus on these relationships, all four modern taxa or OTUs of Chrysemys were included in the dataset, rather than only Chrysemys picta picta Schneider, 1783 that had been included by Jasinski (2018a). Fossil species previously referred to Chrysemys were also included in the current analysis, including ‘ C.’ carri , C. timida and ‘ C.’ aeilliamsi . Chrysemys isoni was not included in the phylogenetic analysis due to the fragmentary nature of its fossil material, as it acted as a wildcard taxon. Phylogenetic analyses were run with molecular constraints on the genera of modern OTUs. Constraints were based on the recent intergeneric relationships recovered by Spinks et al. (2016: fig. 5), namely Deirochelys + ( Chrysemys + ( Pseudemys + (( Graptemys + Malaclemys ) + Trachemys ))). Species were not constrained. This was done to constrain relationships between genera, allow morphological data to determine the interspecific relationships within genera, and to determine the generic placement and phylogenetic relationships of the included fossil species.
Maximum parsimony analyses were run using TNT v.1.5 ( Goloboff & Catalano, 2016). A total of 36 taxa were analysed, including 35 deirochelyines (ingroup taxa). Clemmys guttata (Schneider, 1792) was used as the outgroup as it has been found to be a basal member of the sister subfamily Emydinae , particularly with morphological data (e.g. Gaffney & Meylan, 1988; Stephens & Wiens, 2003). A total of 246 characters were scored, with 96 cranial (39% of total), 23 post-cranial (9%), 82 directly from the osteology of the shell (33%) and 45 from the scutes of the shell (19%). All characters were left unweighted, a branch-and-bound search was used with minimum branch lengths set to collapse. This resulted in 25 most parsimonious trees with a consistency index (CI) of 0.413, a retention index (RI) of 0.515, and a tree length of 881 steps. Both the strict consensus ( Fig. 8 View Figure 8 ) and 50% majority rule consensus trees ( Fig. 9 View Figure 9 ) are reported. The 50% majority rule consensus tree resulted in more resolution in some areas of the tree (e.g. Graptemys species, Deirochelys species and Chrysemys species). Hypotheses of the phylogenetic relationships are discussed based on findings in the present study and comparing these with previous studies.
DISCUSSION
Chrysemys corniculata is identified as a member of the genus Chrysemys by several features discussed above in the diagnosis, including; a low domed shell, an absent or very weak anal notch, absence of notching between posterior peripherals and marginals, absence of a dorsal keel, vertebral 1 with relatively slight nuchal overlap, and presence of anterolateral flaring/projections of the nuchal. This resulted in Chrysemys , and its presumed relationships, as the focus of the analysis. In addition to C. picta , several modern members of the other deirochelyine genera are included, along with several presumed fossil members of the subfamily. This was done in a similar way to Jasinski (2018a), and the relationships of several taxa from their study are re-evaluated. Chrysemys species, and other deirochelyine taxa, are analysed, or re-analysed, and their evolutionary relationships are reassessed, in particular in regard to their generic placement and species interrelationships.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.