
Dyckia tomentosa Mez (1896: 515)
|
publication ID |
https://doi.org/10.11646/phytotaxa.595.2.4 |
|
DOI |
https://doi.org/10.5281/zenodo.7908732 |
|
persistent identifier |
https://treatment.plazi.org/id/FB6AEA65-FF98-FFC2-22F0-FB0CE778FA2B |
|
treatment provided by |
Plazi |
|
scientific name |
Dyckia tomentosa Mez (1896: 515) |
| status |
|
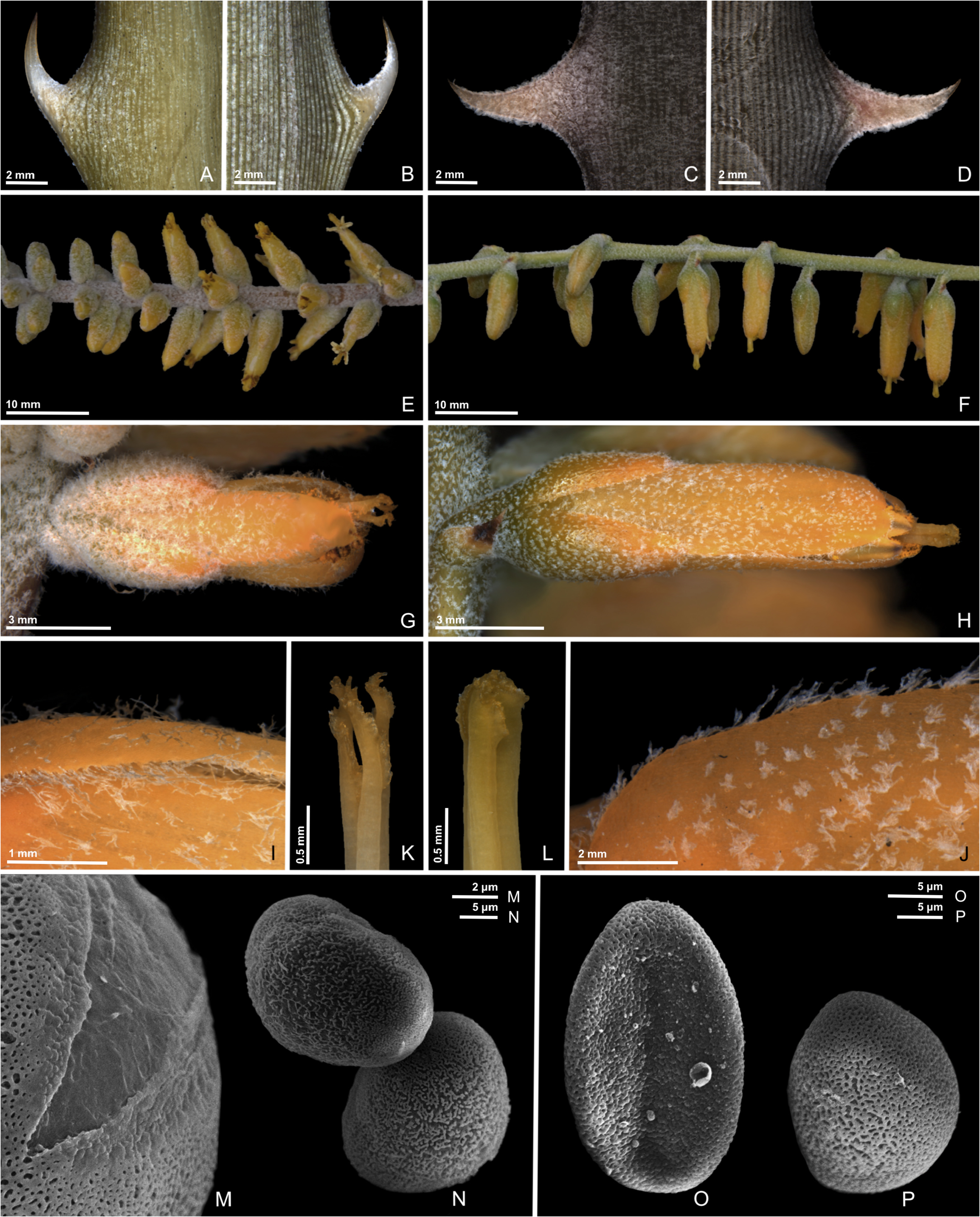
2. Dyckia tomentosa Mez (1896: 515) View in CoL . Figures 2A, B, E, G, I View FIGURE 2 , and K.
Type: — BRAZIL. [ Rio Grande do Sul: Taquari], “ Co d’Utra” [Cerro Dutra], 1 September 1823, F. Sellow s.n. ( holotype: P00713136 [ P image!]; isotype [fragment]: B _10_0248899 [ B image!]).
= Dyckia polyclada Smith (1989: 206) , syn.nov.
Type: — BRAZIL. “Almost certainly Rio Grande do Sul ”, April 1988, R. Foster 3096 ( holotype: US00169775 and US 00169776 [US image!]; isotypes: GH00067261 [ GH image!], NY00335407 [ NY image!], P00105589 [ P image!]).
Herb saxicolous, rhizomatous, 1.5–2.3 m in height when in bloom; rosette symmetrical or slightly asymmetrical, (29–) 46–190 cm in diameter. Leaves ca. 60 in number, the inner ones erect, the outer ones patent to reflexed; blades (13–)22–96 × 1.6–8 cm, narrowly triangular, rigid, adaxially green to slightly cinereous, sparsely to densely lepidote, trichomes adpressed, abaxially longitudinally nerved-striate, green to cinereous, lepidote between the nerves, margins spinose-serrate; spines 2–7 × 0.9–3.8 mm, conspicuous, narrowly triangular, aduncate, antrorse, rigid, whitish-green at the base and yellowish brown at the apex. Inflorescence axillary, erect; peduncle 38–87 cm long, robust, reddish green to completely red, glabrescent, pruinose; peduncle bracts of the median portion polystichous, stramineous, equaling or surpassing the internodes; fertile part of the inflorescence 74–138 cm long, twice-branched; primary bracts identical to the peduncle bracts; first order branches 9–62 in number, suberect to patent, with reddish green to red axes, glabrescent, pruinose and with white trichomes; second order branches when present usually less than 3, rarely 4–5, at the base of the first order branches; floral bracts 3–4 × ca. 1.5 mm, ovate to lanceolate, the basal portion green, the median portion stramineous and yellowish, the apex stramineous and yellowish brown, attenuate-acuminate, slightly carinate or ecarinate, tomentose, exceeded by the sepals. Flowers 0.8–1.2 cm long, patent to suberect, inconspicuously pedicellate to sessile; pedicels cylindrical, up to 0.5 mm long, greenish, tomentose; sepals 4–5 × ca. 2 mm, triangular-ovate to triangular-lanceolate, apex obtuse to rounded, yellowish green, slightly carinate, abaxial surface densely tomentose, covered by dendritic trichomes, the margins with filiform uniseriate or slightly branched trichomes; dendritic trichomes on the sepal abaxial surface with an uniseriate stalk of (2)3–4(5) cells, region of branching somewhat expanded, with 3–7 branches formed by 3–4 elongated cells; filiform trichomes of the sepal margins uniseriate or little branched with two branches at most, 4–9 cells; petals 6–7.6 × ca. 2 mm, not unguiculate, erect, oblong to oblanceolate, the basal portion greenish, the median to apical portion yellowish, apex rounded or emarginate, abaxial surface tomentose, covered by dendritic trichomes, the margins with filiform uniseriate trichomes; dendritic trichomes on the petal abaxial surface identical to the trichomes of the sepals surfaces, although more sparsely arranged; filiform trichomes of the petal margins identical to those of the sepal margins, although bigger, 8–16 cells; stamens included during theca opening, subsequently becoming generally exserted afterward; filaments 4–6 mm long, free above the hypanthium; pistil 7–9 mm long; ovary about ca. 1.5 × 1.3 cm; style 4–5 mm long; stigma exserted at anthesis, stigmatic lobes conduplicate, slightly spirally twisted at pre-anthesis, subsequently becoming erect. Capsules ovoid, dark brown when mature. Seeds oblanceoloid.
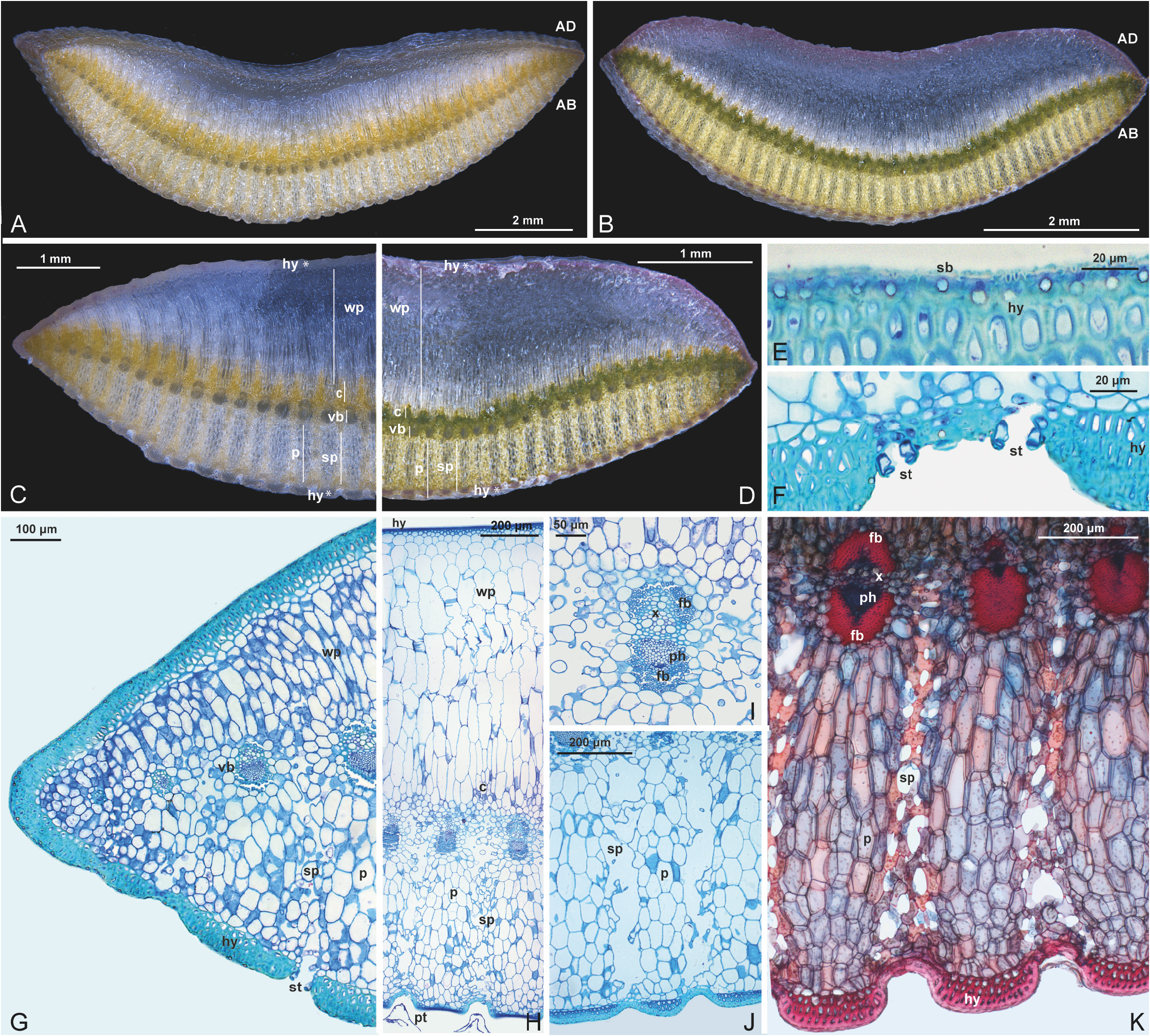
Leaf anatomy: —The analysis of the leaf anatomy of D. tomentosa enabled the identification of the following tissues: epidermis, water-storage parenchyma, chlorenchyma, vascular bundles, spongy parenchyma, ground parenchyma and mechanical hypodermis ( Fig. 3A and 3C View FIGURE 3 ). The epidermis consists of cells that contain silica bodies and present a more pronounced thickening of the anticlinal and internal periclinal walls, which reduces the cell lumen ( Fig. 3 E View FIGURE 3 ). The stomatal complexes are located on the abaxial surface of the leaf blades upward the level of the epidermal cells. The substomatal chambers are associated with the spongy parenchyma, which has long arms and provides a larger dimension of the area of these chambers and is located between the vascular bundles. The mesophyll is presented on the abaxial and adaxial surfaces, a mechanical hypodermis rich in lignin, consisting of three cell layers ( Fig. 3H View FIGURE 3 ). On the adaxial surface it is possible to identify water-storage parenchyma composed of two or three layers of isodiametric cells and subsequently four to five layers of anticlinally elongated cells, with ca. 1.5 mm height ( Fig. 3H View FIGURE 3 ). This parenchyma extends to the region close to the vascular bundles, where it is interspersed with projections of chlorenchyma, 150–200 μm in height. The bundle sheath is continuous around the vascular bundle. The collateral vascular bundles are linearly distributed throughout the leaf section and have lignified sclerenchymatic fibers at the pole of the xylem and of the phloem ( Fig. 3K View FIGURE 3 ). The diameter of the vascular bundles varies along the cross section of the leaf, with larger caliber bundles having fiber caps of the same thickness, both at the xylem pole and at the phloem pole ( Fig. 3K View FIGURE 3 ). The smaller caliber bundles have thicker sclerenchyma fibers at the phloem pole. On the abaxial surface, it is possible to identify spongy parenchyma interspersed with ground parenchyma ( Fig. 3J–K View FIGURE 3 ). The spongy parenchyma, measuring 800–1000 μm height × 100–200 μm width, begins in the region between the vascular bundles and extends to the region of the epidermis, where the stomata and trichomes are located ( Fig. 3J View FIGURE 3 ). The ground parenchyma ( Fig. 3J View FIGURE 3 ), measuring 700–800 μm height × 200–250 μm width, begins in the region of the vascular bundles and extends to the region of the mechanical hypodermis ( Fig. 3K View FIGURE 3 ).
Pollen morphology: —The pollen grains of D. tomentosa are oblate, heteropolar, medium in size (25–50 µm) ( Hesse et al. 2009), and sulcate of the simple sulcus type ( Halbritter & Hesse 1993) ( Fig. 2M and 2N View FIGURE 2 ). The ornamentation has a reticulate to micro-reticulate and heterobrochate pattern ( Punt et al. 2007). However, this comprehensive nomenclature is not entirely adequate for the variations observed. Several discontinuities can be observed in the reticulum roof, in a generalized way ( Fig. 2N View FIGURE 2 ). This pattern has no correspondence in any type already described. In addition, its characterization as semitectate complements the description of ornamentation ( Punt et al. 2007). Wanderley & Melhem (1991) described discontinuities in the pollen reticulum of Dyckia species, however with less intensity. The discontinuity observed in D. tomentosa is widespread, similarly to that observed in Pitcairnia albiflos Herbert (1826 : t. 2642) ( Halbritter 2016).
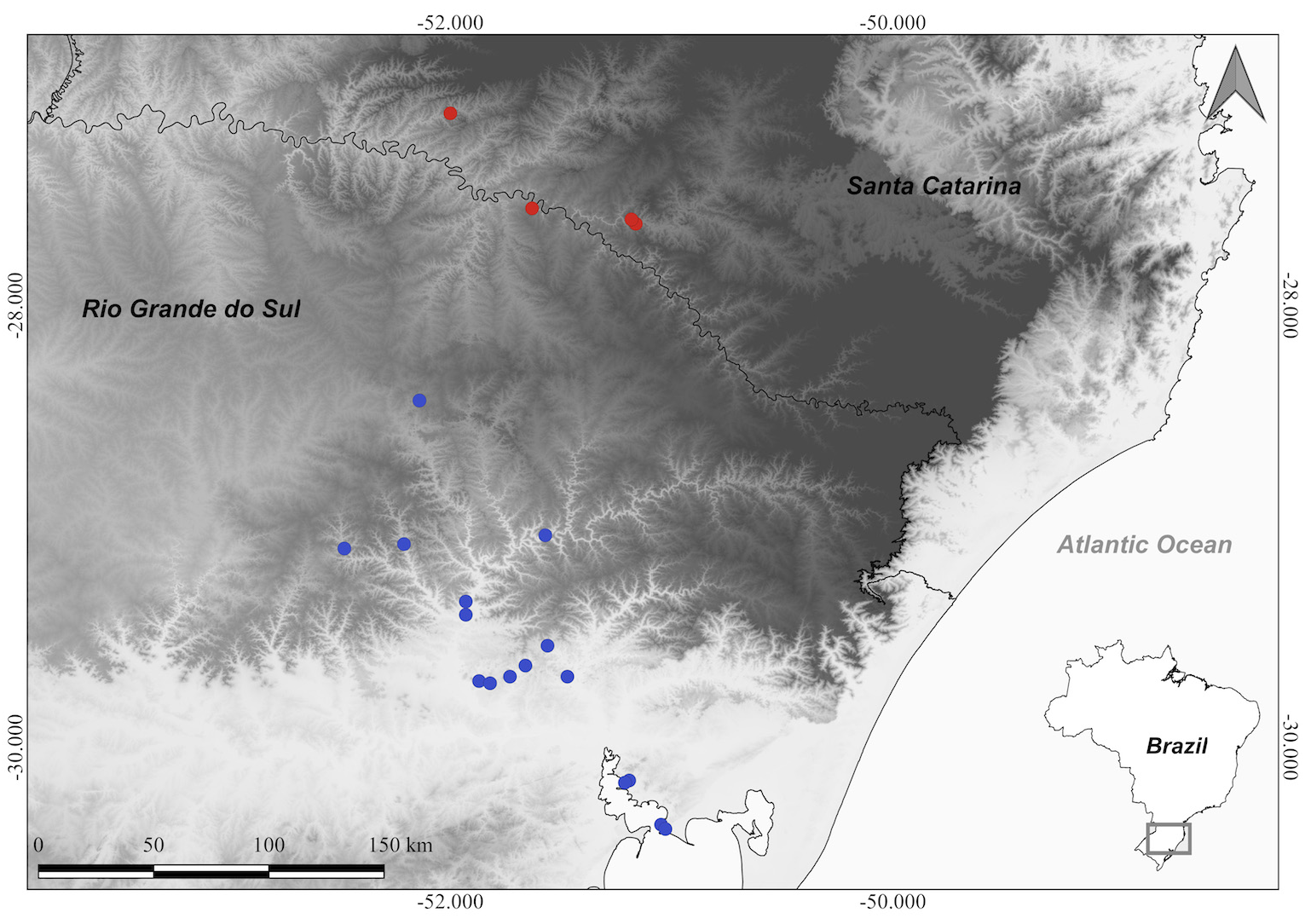
Distribution & habitat: —Occurs as a saxicolous heliophyte on rocky outcrops or rocky escarpments of basalt, granite or sandstone. Most of the known populations are found at low altitudes (< 100 m) in the hydrographic basins of the Taquari and Jacuí rivers, and Guaíba lake, in extreme southern Brazil, in the state of Rio Grande do Sul ( Fig. 4 View FIGURE 4 ). Most of the populations have a significant number of individuals (>200), however, some have been suppressed by urban advancement, as they occur in the most densely populated regions of the state of Rio Grande do Sul, close to the state capital.
Observations:— Dyckia tomentosa has been neglected for decades despite being one of the most common and collected species in the D. selloa complex. Since its description by Mez (1896), there have been doubts about the identity of the species since its holotype (the only known sample of the species that its author had access to, deposited in herbarium P, with a fragment in herbarium B), consisted of only a piece of a leaf and the apical part of a degraded already-fruiting inflorescence (the fragment in B is only a piece of the branch of the fruiting inflorescence). The label attached to the plant by the naturalist Friedrich Sellow was enigmatic, and it was only after an extensive study of Sellow (by the first author of this article), including a study of his handwriting, that it was possible to decipher the information on the label. This label contained a rare record of information, consisting of the date (“1 spter 23” = 1 September 1823) and location of collection (“ Co d’Utra” = Cerro Dutra) (Sellow did not usually include locations and dates on the labels of plant specimens, but only the collection number). This information cross-referenced with the literature related to the naturalist ( i.e. Urban 1893; Weiss 1941; Herter 1945; Marchiori et al. 2018) led to the discovery of the exact collection locality, place known today as “Morro do Carapuça”, a rocky escarpment from a hill in the north of the city of Taquari ( Rio Grande do Sul) whose geology and physiognomy are described in the text of Weiss (1941) under the old name “Cerro d’Utra”. Thus, we were able to collect the species again at its type locality after almost 200 years and definitively establish its identity. The type population currently has ca. 30 individuals on a rocky escarpment.
The identity of the species, due to its poor type specimen, was in doubt by Smith (1956),and later it was synonymized by Smith (1966) under D. maritima . It was kept as a synonym until B̧neker et al. (2020; 2021) re-established it without further explanation. The explanations are given in this work. Smith (1989) described D. polyclada (originally described with the incorrect grammatical gender “ D. polycladus ”) based on material of dubious origin. However, this name was little applied later and mistakenly applied to D. selloa by Strehl (1998). From our observations of the populational morphological variation of the species, we conclude that the most likely origin of the specimen described as D. polyclada , is “Serra Gaúcha”. We consider what was described as D. polyclada in an ecotype, characterized by being more robust than the specimens of most other populations of D. tomentosa . The population that occurs in the municipality of Bento Gonçalves ( H.M. B¸neker 398 et al. (HDCF)) seems to be the closest to the type of D. polyclada , and revealed itself a synonym of D. tomentosa from careful analysis of fertile material. Dyckia maritima (known only from the cliffs of the municipality of Torres, Rio Grande do Sul, Brazil) differs greatly from D. tomentosa , and does not seem to be so closely related, except for the presence of indumentum on the axes of the inflorescence, sepals and petals. When revising Prionophyllum, Mez (1935) considered only two species, and synonymizing several other names, whose differentiation criterion was basically the presence ( P. maritimum ( Baker 1889: 136) Mez (1896: 542)) or absence ( P. selloum ) of indumentum in the inflorescence, a treatment followed by Smith & Downs (1974). Dyckia tomentosa is, in fact, morphologically very similar to D. hebdingii , differing basically by its larger size when flowering (1.5–2.3 vs. 0.4–1 m high.), larger inflorescence (up to 2.1 vs. up to 0.95 m long) with more branches (9–62 vs. 3–12 branches), and the direction of the stigmatic lobes at anthesis (erect vs. spirally twisted). Dyckia magnifica , described here, is also similar to D. tomentosa , with their distinctions discussed below.
Specimen examined:— BRAZIL. Rio Grande do Sul: Arroio do Meio, Morro Gaúcho , 1 November 1991, J. A. Jarenkow 1933 ( PEL); 14 October 2012, H. M. B ¸neker 109 & A. Schossler ( HDCF) ; Barros Cassal, perto da ponte caída do rio F „o, 22 January 2014, H. M. B ¸neker 394 et al. ( HDCF); Bento Gonçalves, 17 October 1988, N. Silveira 8803 ( HAS); 4 November 1982, J. Mattos & N. Mattos s.n. ( HAS 66151 About HAS ); 3 November 1995, T. Strehl 1429 ( HAS); 9 February 2015, H. M. B ¸neker 398 et al. ( HDCF); Maratá, Cascata Vitória , 8 June 2015, H. M. B ¸neker et al. 490 et al. ( HDCF); Marau, Cachoeir „o, 21 February 2015, H. M. B ¸neker 419 et al. (HDCF); Montenegro, Morro do Cabrito , 8 January 1986, I. Fernandes 40 ( ICN); 30 November 1988, I. Fernandes 419 ( ICN); morro na cidade, 10 February 2015, H. M . Bu ̈neker 405 et al. ( HDCF) ; Porto Alegre, Morro Tapera , 8 May 1979, O. Bueno 1372 ( HAS); Morro das Abertas , 31 October 2015, H. M. B ¸neker 495 et al. (HDCF); São José do Herval, PCH Salto do Forqueta , 27 January 2003, E. Musskopf s.n. ( HVAT 1109 ); Tabaí, 30 January 2014, H. M. B ¸neker 251 et al. (HDCF); H.M. B¸neker 253 et al. ( HDCF); 21 January 2017, H. M. B ¸neker 609 ( HDCF); 10 January 2017, C. J . Breitsameter s.n. et al. ( ICN 188086 View Materials ) ; Taquari, Cerro Pelado, 4 January 1996, s.c. ( ICN 111080 View Materials ); Morro do Carapuça ( Cerro Dutra ), 21 January 2017, H. M. B ¸neker 607 et al. (HDCF); Teutônia, Lagoa da Harmonia , 25 December 2017, H. M. B ¸neker 700 (HDCF); Viamão, Itapu „, 15 Mach 1994, T. Strehl 1268 ( HAS); s.d., T. Strehl 1269 ( HAS); s.d., T. Strehl 1270 ( HAS); October 1986, Lúcia & T. Strehl 1342 ( HAS); 28 Mach 1996, C. C. Waldemar s.n. ( ICN 111516 View Materials ); Morro do Araçá , 14 January 2003, M. Pinheiro 421 ( ICN); Morro da Grota, 31 January 1975, M. L. Porto s.n. et al. ( HAS 1175 About HAS ).
| F |
Field Museum of Natural History, Botany Department |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| R |
Departamento de Geologia, Universidad de Chile |
| GH |
Harvard University - Gray Herbarium |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| J |
University of the Witwatersrand |
| A |
Harvard University - Arnold Arboretum |
| PEL |
Universidade Federal de Pelotas |
| H |
University of Helsinki |
| M |
Botanische Staatssammlung München |
| N |
Nanjing University |
| HAS |
Fundação Zoobotânica do Rio Grande do Sul |
| T |
Tavera, Department of Geology and Geophysics |
| I |
"Alexandru Ioan Cuza" University |
| ICN |
Instituto de Ciencias Naturales, Museo de Historia Natural |
| O |
Botanical Museum - University of Oslo |
| PCH |
Prestwich and Pilkington Botanical Society |
| E |
Royal Botanic Garden Edinburgh |
| C |
University of Copenhagen |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dyckia tomentosa Mez (1896: 515)
| Büneker, Henrique Mallmann, Pastori, Tamara, Almeida, Pedro Schwambach De & Mariath, Jorge Ernesto De Araujo 2023 |
Dyckia polyclada
| Smith, L. B. 1989: ) |