
Araneibatrus Yin and Li, 2010
|
publication ID |
https://doi.org/10.11646/zootaxa.4097.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:09889BBB-66DE-46D2-A394-6A194E7AE102 |
|
persistent identifier |
https://treatment.plazi.org/id/F340E468-FF81-FFF5-5783-86E6FEFFF9BE |
|
treatment provided by |
Plazi |
|
scientific name |
Araneibatrus Yin and Li, 2010 |
| status |
|
Araneibatrus Yin and Li, 2010 View in CoL
Araneibatrus Yin & Li, 2010 View in CoL (in: Yin, Li & Zhao 2010: 54). Type species: Araneibatrus gracilipes Yin & Li, 2010 View in CoL (original designation).
Tribasoadellus Nomura & Yin, 2011 (in: Yin, Nomura & Zhao 2011: 37), new synonymy. Type species: Batrisodellus callissimus Nomura & Wang, 1991 View in CoL (original designation).
Revised diagnosis. Body, antennae, and legs distinctly elongate. Head roundly rectangular; vertex with small, nude foveae situated posterior to dorsal level of posterior margin of eyes; lacking frontal rostrum; with strongly prominent antennal tubercles. Pronotum lacking median antebasal fovea (fovea reduced to a pit or an impressed area), lacking lateral longitudinal sulci and marginal spines. Elytra narrowing to bases; each elytron with three basal foveae; lacking distinct discal stria. Abdominal tergite IV (first visible tergite) longest, with oblique inner marginal carinae. Aedeagal median lobe asymmetric, with large basal capsule and foramen, with articulated dorsal lobe.
Redescription. Length 2.01–2.61 mm. Head roundly rectangular; lacking frontal rostrum, with distinctly prominent antennal tubercles; vertexal foveae nude, situated posterior dorsal level of posterior margins of eyes, lacking sulcus connecting foveae; in some species vertex with mediobasal carina, lateral post-antennal carinae present or absent; antenna with eleven antennomeres, clubs formed by three apical antennomeres, in some cases clubs indistinctly delimited; ocular-mandibular carinae present, extending from anterior margins of eyes, across frons to clypeal anterior margin; clypeus with rounded anterior margin, one-fifth to one-third of total head length; eyes prominent in male, posteroventral margins slightly flat; maxillary palpus with palpomere I minute, II elongate and pedunculate at base, III short and triangular, IV fusiform, its mesal margins roundly expanded, with small palpal cone; with single gular fovea.
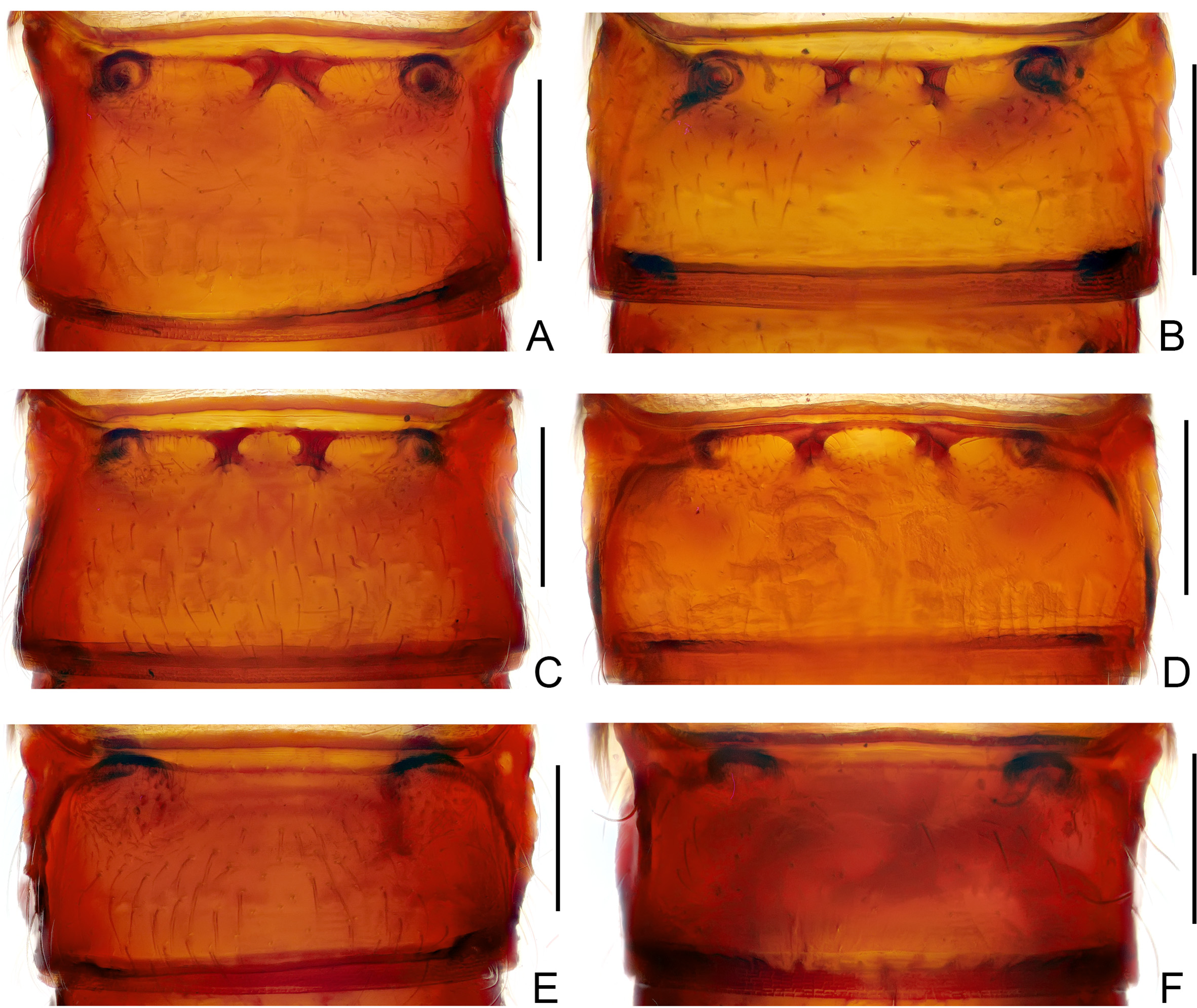
Pronotum lacking median antebasal fovea, lateral antebasal foveae connected by shallow transverse antebasal sulcus, sometimes sulcus faint or lacking; inner and outer pairs of basolateral foveae present; median longitudinal and antebasal sulci distinct to lacking, lacking lateral longitudinal sulci; laterodorsal carinae rarely present; discal or antebasal tubercles/spines rarely present; lateral margins lacking spine; paranotal carinae complete; lateral procoxal foveae present.
Each elytron with three small to large nude foveae at base; lacking subbasal fovea; discal stria indistinct or lacking; lacking subhumeral foveae, marginal stria present in posterior half of elytral lateral margin; apicolateral margins straight to shallowly emarginate.
Mesoventrite with median foveae widely separated, opening into shared transverse cavity, lateral mesoventral foveae forked, often partially overlapping median foveae; lateral mesocoxal foveae large and deep. Metaventrite with lateral foveae; metacoxae widely separated; posterior margin of metaventrite with narrow or wide notch at middle.
Abdominal tergite IV (first visible tergite) as long as to slightly longer than tergites V and VI combined; with complete outer pair, and usually complete and oblique inner pair of marginal carinae; short discal carinae sometimes present; basal impression complete when with only two basolateral foveae, or interrupted when pair of mediobasal and/or two pairs of basolateral foveae present; tergites V and VI combined slightly longer than VII, single lateral carina present or absent; tergites V–VII each with one pair of basolateral foveae; tergite VII with pair of small lateral tubercles (clearly seen only in slide preparation). Sternite IV (second visible sternite) usually with one pair of mediobasal and three pairs of basolateral foveae, occasionally mediobasal foveae lacking; sternites V– VII each short at midline, with one pair of small basolateral foveae.
Legs with pro- and mesotarsomeres II as long as III, metatarsomeres II slightly longer than III.
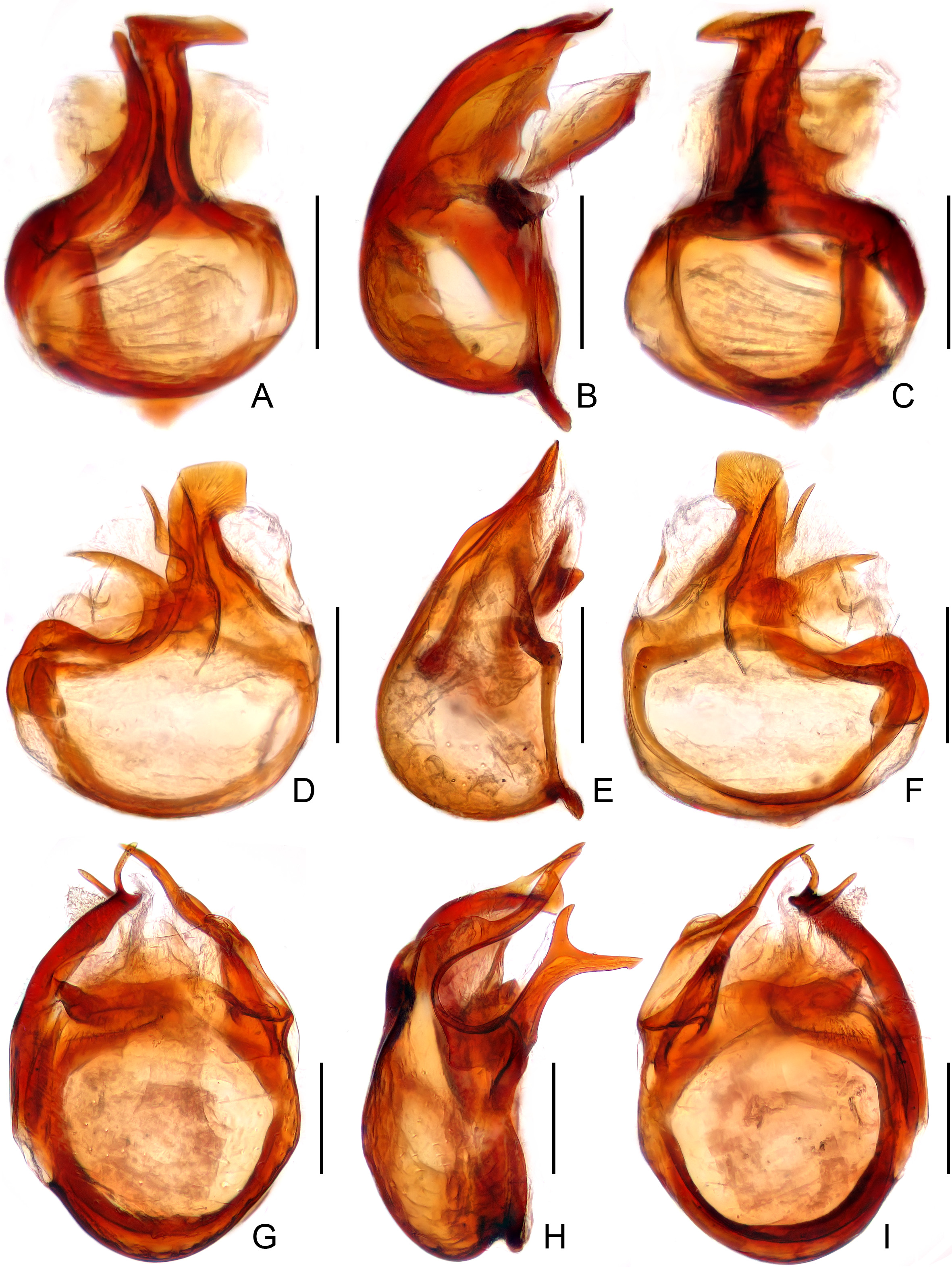
Male secondary sexual characters: larger eyes and longer elytra; antennal clubs and metaventrite modified; mesotrochanter, mesotibia, or metatrochanters spinose. Aedeagus with parameres fused to median lobe, with large basal capsule and foramen, articulated dorsal lobe elongate, in dorsal view offset to left side.
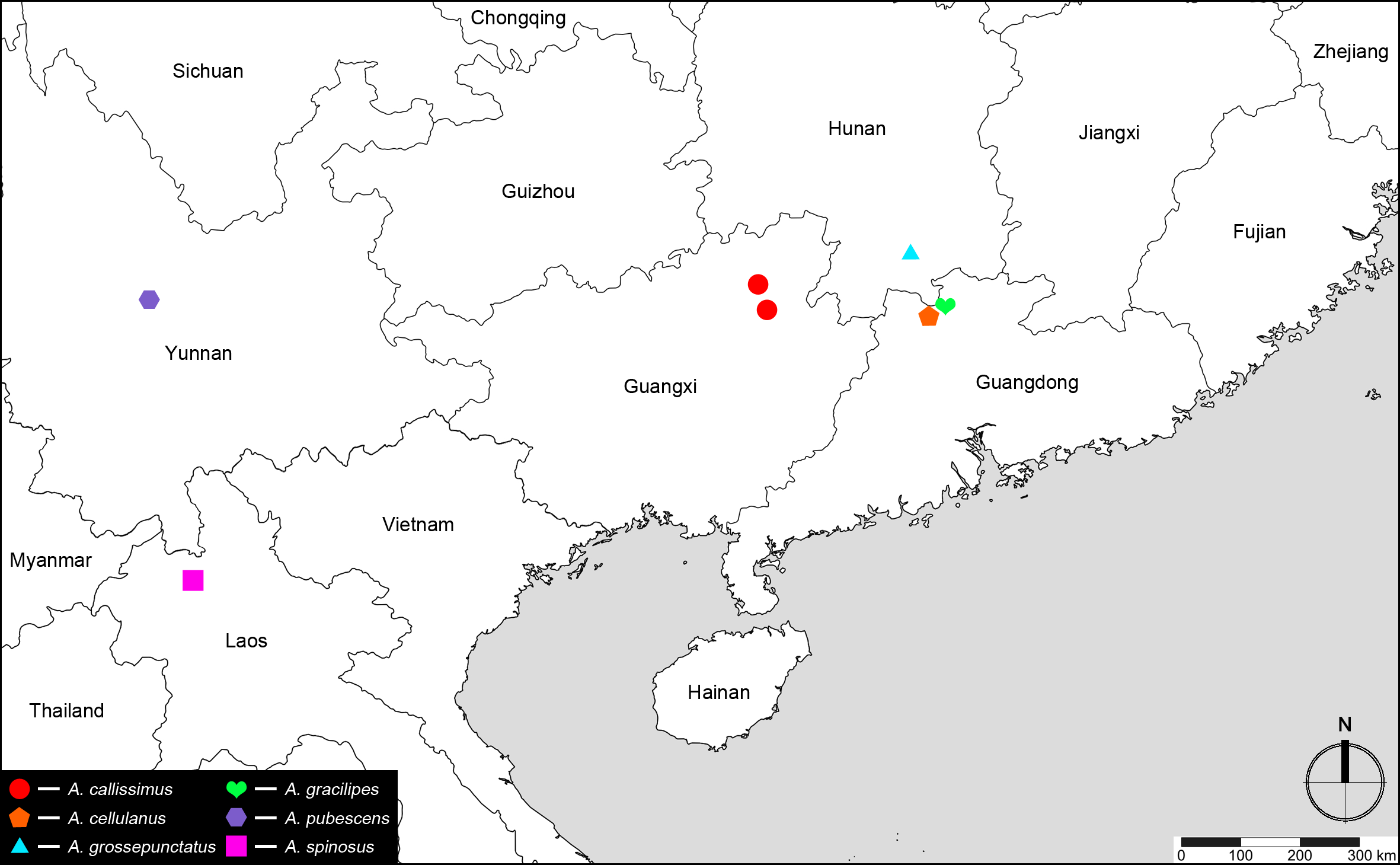
Discussion. Araneibatrus was originally established (Yin, Li & Zhao 2010: 54) based on a single species A. gracilipes collected in leaf litter samples in the Nanling Nature Reserve, southern China. This species possesses relatively long antennae and legs (Yin, Li & Zhao 2010: fig. 1), similar to a few batrisine genera that include cavernicolous species. In the following year, a new generic name Tribasodellus was proposed ( Yin, Nomura & Zhao 2011) for Batrisodellus callissimus , a species know to occur in several limestone caves around the Guilin City, Guangxi ( Nomura & Wang 1991). But Tribasodellus was not compared with any relatives at that time.
Upon a reexamination and comparison of the type species of both genera, we found that Araneibatrus differs from Tribasodellus mainly in the following characters states: 1) the head, pronotum, and elytra much more slender in Tribasodellus than those of Araneibatrus; 2) presence of a pair of lateral post-antennal carinae, and a short median carina on the vertex in Tribasodellus, both carinae absent in Araneibatrus; 3) the pronotum with paired discal and antebasal tubercles in Araneibatrus, the tubercles absent in Tribasodellus; 4) the basolateral margins of the elytra shallowly emarginate (the elytral base more constricted) in Tribasodellus, the basolateral margins not emarginate in Araneibatrus; 5) the lateral and median mesoventral foveae largely overlapping in Tribasodellus, the foveae clearly separated in Araneibatrus; 6) the posterior margin of the metaventrite at the middle with a wide notch in Tribasodellus, the notch narrow in Araneibatrus; 7) the tergite IV with only two pairs of basolateral foveae in Tribasodellus, with one pair of mediobasal and two pairs of basolateral foveae in Araneibatrus; and 8) the aedeagal median lobe with a wide, transverse basal capsule in Tribasodellus, the basal capsule large and slightly elongate in Araneibatrus. The question is whether these differences are sufficient to justify the placement of Araneibatrus and Tribasodellus as separate genera.
Newly acquired material from various collections was examined to clarify this problem. Two new species collected in caves from central and southern China, described as A. cellulanus sp. n. and A. grossepunctatus sp. n. below, are clearly congeneric with T. callissimus by sharing the numerous character states that define the genus. The new species possess a distinctly elongate body ( Figs 1 View FIGURE 1 B, 2A), the vertex with a pair of postantennal lateral carinae, and the pronotum lacking any tubercles or spines ( Fig. 4 View FIGURE 4 B–C), as is found in in T. callissimus . On the other hand, both new species have one pair of mediobasal and two pairs of basolateral foveae at the base of tergite IV ( Fig. 8 View FIGURE 8 B–C), same as that in A. gracilipes ( Fig. 8 View FIGURE 8 D). In A. cellulanus , the median notch at the posterior margin of the metaventrite is slightly narrower ( Fig. 6 View FIGURE 6 B) than that in T. callissimus ( Fig. 6 View FIGURE 6 A), and in A. grossepunctatus this notch is yet narrower ( Fig. 6 View FIGURE 6 C). Araneibatrus grossepunctatus additionally has a pair of dorsal carinae on the pronotal disc ( Fig. 4 View FIGURE 4 C), which are absent both in T. callissimus and A. gracilipes . The aedeagal form of A. cellulanus ( Fig. 10 View FIGURE 10 D–F) is similar to that of T. callissimus ( Fig. 10 View FIGURE 10 A–C) by sharing a transverse basal capsule, while the aedeagal basal capsule of A. grossepunctatus ( Fig. 10 View FIGURE 10 G–I) is much stouter.
Another two new species from southwestern China and northern Laos, described below as A. pubescens sp. n. and A. spinosus sp. n., have an extremely elongated body form, antennae, and legs ( Fig. 3 View FIGURE 3 A–B). They obviously belong in the same genus and seem to be congeneric with T. callissimus by sharing a similar foveal pattern and aedeagal form. These two new species lack lateral and mediobasal carinae on the vertex ( Fig. 4 View FIGURE 4 E–F), and have a narrow median notch of the metaventrite ( Fig. 7 View FIGURE 7 A–B), similar to the condition known in A. gracilipes . However, the new species do not have the middle longitudinal and antebasal sulci on the pronotum ( Fig. 4 View FIGURE 4 E–F), and have a reduced number of basolateral foveae (only two) on tergite IV ( Fig. 8 View FIGURE 8 E, 8F). In A. spinosus , the pronotum has a pair of large antebasal spines ( Fig. 4 View FIGURE 4 F), and sternite IV uniquely lacks mediobasal foveae ( Fig. 9 View FIGURE 9 F). The aedeagal basal capsule of A. pubescens ( Fig. 11 View FIGURE 11 D–F) is distinctly transverse, similar to that of T. callissimus and A. cellulanus , while in A. spinosus it is quite stout ( Fig. 11 View FIGURE 11 G–I), similar to that of A. grossepunctatus .
It can be concluded for Tribasodellus and Araneibatrus that 1) the body form varies among the species that live in different types of habitat, with cavernicolous species usually having a more slender body, antennae, and legs; 2) the lateral and mediobasal carinae of the vertex, the pronotal discal and antebasal tubercles/spines, as well as the pronotal median longitudinal and transverse antebasal sulci can be present or absent; 3) the species that lives in leaf litter ( A. gracilipes ) has a complete set of the abdominal foveae, with six basal foveae on the tergite IV and eight on the sternite IV, while in cavernicolous species, the number of basal foveae can be reduced to two to four on tergite IV and six on sternite IV; and 4) the aedeagal basal capsule is most elongate in A. gracilipes , and stout to distinctly transverse in the cavernicolous species.
Reduction of structures and trend for elongation of the body can be found in other genera of cavernicolous/leaf litter-dwelling Batrisini. Recently ten new cavernicolous Tribasodites species were described from southern China and northern Laos ( Yin, Nomura & Li 2015), whose placement in one genus is strongly supported by the shared uniquely modified male metaventrite, and structure of the aedeagus. Among them, three species lack spines on the lateral margins of the pronotum, whereas the presence/absence of pronotal lateral spines is usually a stable and reliable character to define a genus. In the American species of Batrisodes Reitter , the surface/leaf litter dwellers have a stouter habitus (and with more eye facets), and various grades of becoming elongate (and fewer to no eye facets) are found in the cavernicolous species (Chandler per. comm.).
As there are no characters that can be used to consistently separate Tribasodellus and Araneibatrus, these two genera are synonymized here, with Araneibatrus being the senior synonym, syn. n. Consequently, the only member of Tribasodellus, T. callissimus , is placed in Araneibatrus, resulting in Araneibatrus callissimus comb. n. At present, we have failed to find any synapomorphy to support the monophyly of this derived genus containing both leaf litter-dwelling and cavernicolous species that exhibit different degrees of adaptation, but the combination of the characters listed in the ‘Diagnosis’ section seems to be unique. Pending discovery of new evidence, all new species described in this paper are placed in Araneibatrus.
Comparative notes. Including the new species described in this paper, Araneibatrus now contains one epigean and five cavernicolous species scattered in the southern provinces of China and northern Laos ( Fig. 12 View FIGURE 12 ). The genus is similar to Tribasodites Jeannel containing nineteen species (including thirteen cavernicolous species placed in the ‘ T. bedosae group’) from India, Japan, eastern and southern China, Taiwan, and northern Thailand ( Jeannel 1960; Zhao, Yin & Li 2010; Yin, Li & Zhao 2011; Yin, Nomura & Li 2015). These two genera can be separated primarily by the combination of the more elongate head, pronotum, and elytra, lack of spines on the pronotal lateral margins, and often lack of the pronotal dorsal carinae in Araneibatrus, while members of Tribasodites possess a shorter head, their pronotum is much stouter, often with distinct dorsal discal carinae and marginal spines; also the forms of the antennal modifications and the aedeagal structures are different. Another recently described troglobitic genus Zopherobatrus Yin & Li, with a single species Z. tianmingyii Yin & Li, also exhibits strong cavernicolous adaptations (Yin & Li 2015). It can be readily separated from Araneibatrus by the presence of an anteriorly facing U-shaped sulcus that connects the vertexal foveae, presence of a pair of lateral longitudinal sulci on the pronotum, each elytron with two basal foveae, and aedeagus with a movable articulated dorsal lobe.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pselaphinae |
Araneibatrus Yin and Li, 2010
| Yin, Zi-Wei, Jiang, Ri-Xin & Steiner, Helmut 2016 |
Araneibatrus gracilipes
| Yin & Li 2010 |
Batrisodellus callissimus
| Nomura & Wang 1991 |