
Stenaleyrodes papillote, Martin, Jon H & Streito, Jean-Claude, 2003
|
publication ID |
https://doi.org/10.5281/zenodo.156520 |
|
DOI |
https://doi.org/10.5281/zenodo.6273644 |
|
persistent identifier |
https://treatment.plazi.org/id/ED4E880D-8D2E-FF86-FEBF-FA90D3E5FDFC |
|
treatment provided by |
Plazi |
|
scientific name |
Stenaleyrodes papillote |
| status |
sp. nov. |
Stenaleyrodes papillote View in CoL sp. nov.
( Figs 16 View FIGURE 1 View FIGURE 2. S View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 7 View FIGURE 7 A, 8A, 9, 10)
PUPARIUM ( Figs 14 View FIGURE 1 View FIGURE 2. S View FIGURE 3 View FIGURE 4 , 5 View FIGURE 5 A, 6B, 7A, 10C)
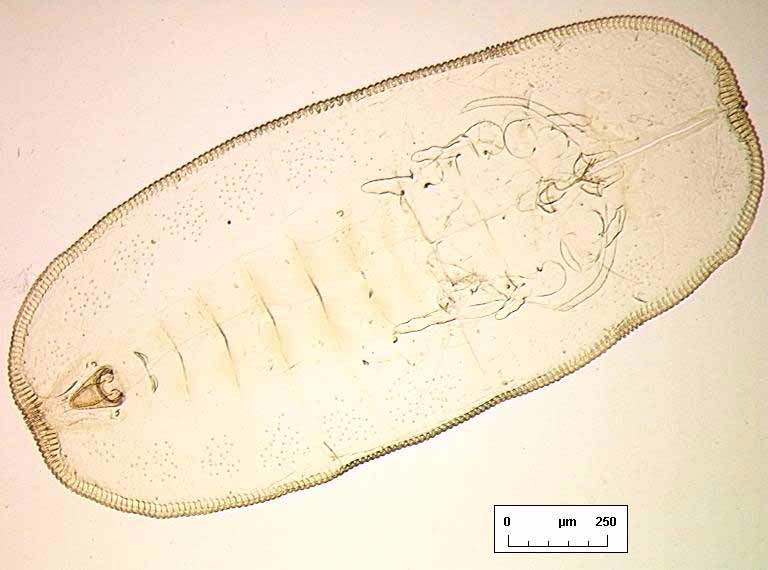
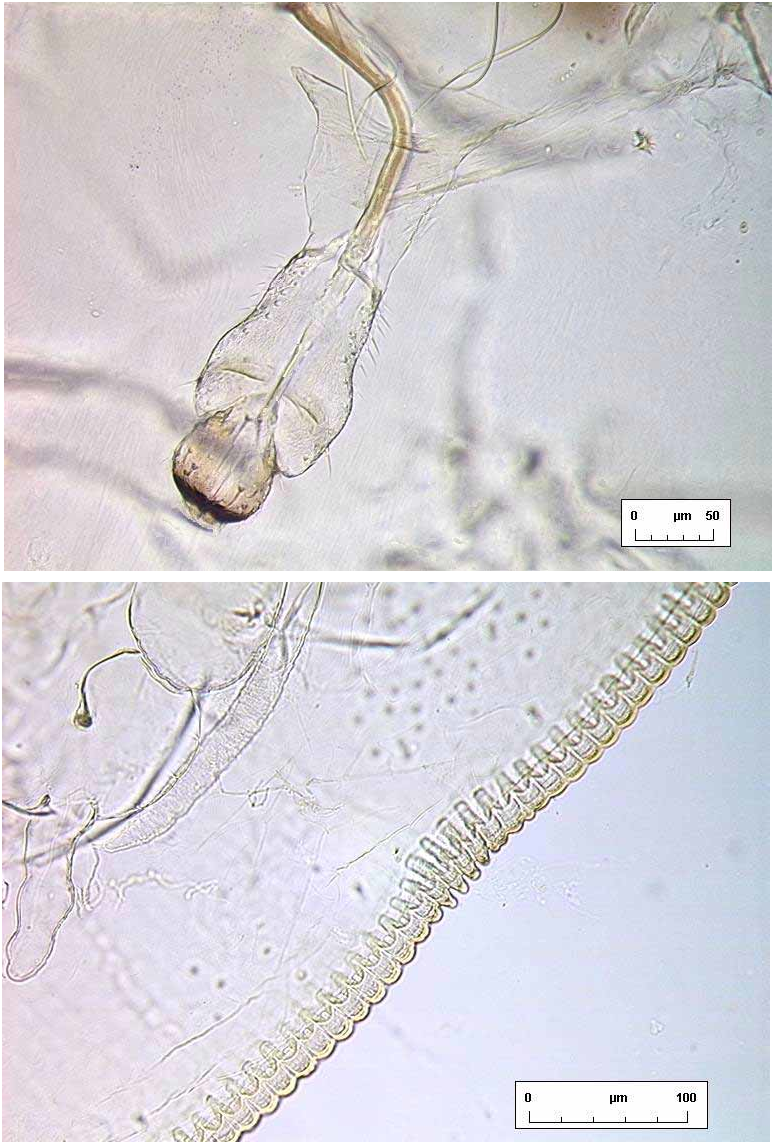
Habitus. When not damaged by abrasion, individual puparia have highly characteristic secretions ( Fig. 1 View FIGURE 1 ) in the form of a very broad, laterally directed, white fringe on each side of the body, and an entirely separate fibrous “brush” of wax at each end, all the secretions in the same plane as the puparium; dorsum with no visible secretions, smooth.
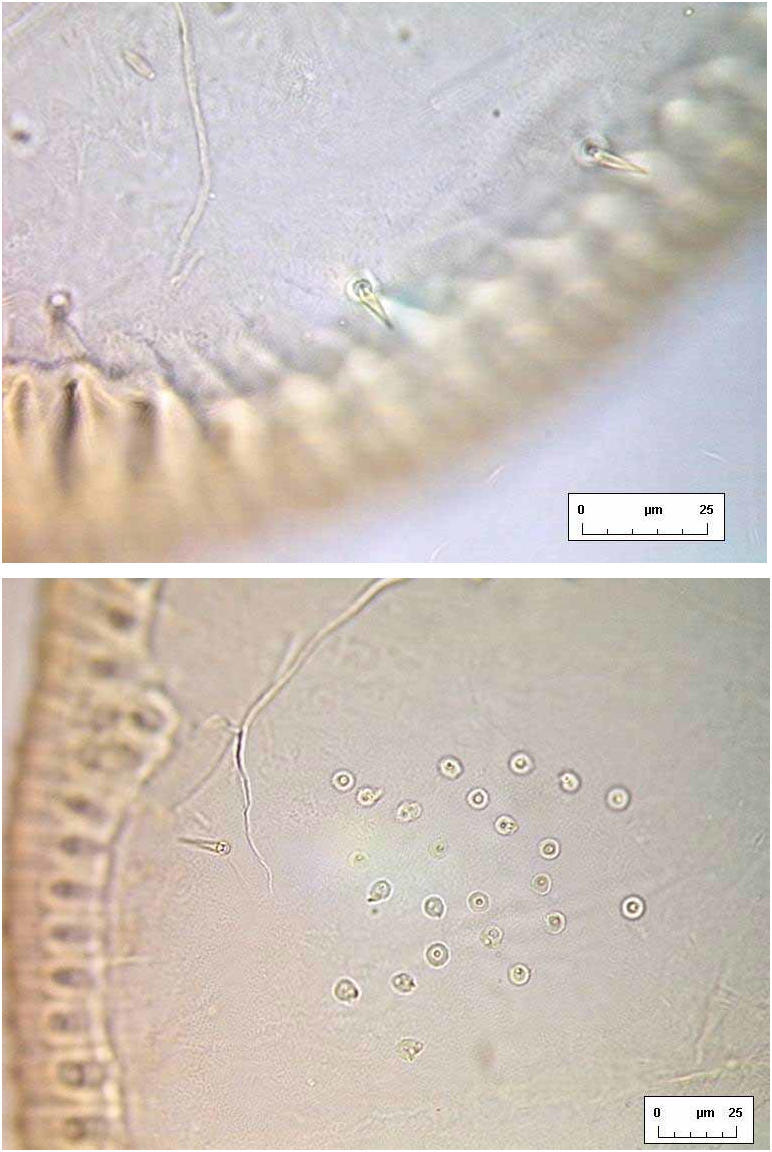
Margin. Outline elongate oval, rather flattened at anterior and posterior ends, approximately twice as long as wide ( Fig. 2 View FIGURE 2. S ). Sexually dimorphic: female puparia 1.882.09 mm long, 0.881.05 mm wide, generally widest at abdominal segment III/IV (n=16); male puparia 1.501.67 mm long, 0.680.76 mm wide, generally widest opposite transverse moulting sutures (n=7). Margin evenly crenulate, with 68 rounded teeth per 0.1mm of lateral margin, and a few teeth are more pronounced at each thoracic ( Fig. 6 View FIGURE 6 B) and caudal tracheal opening.
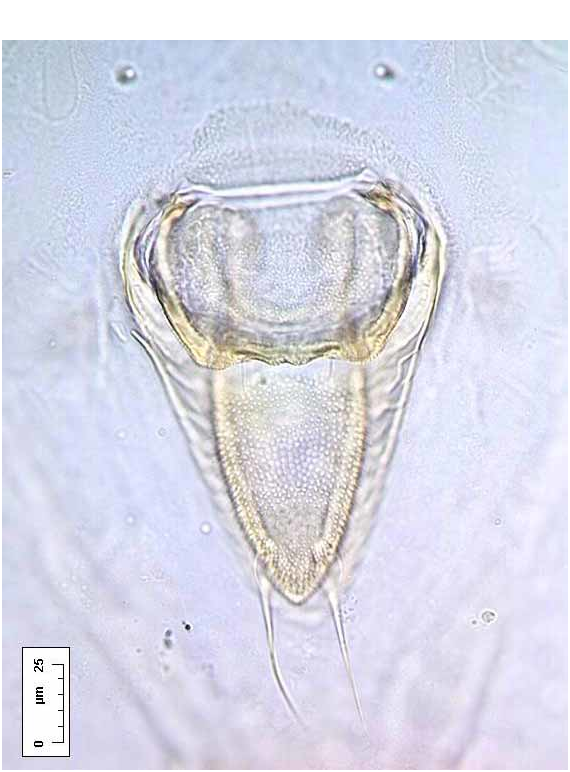
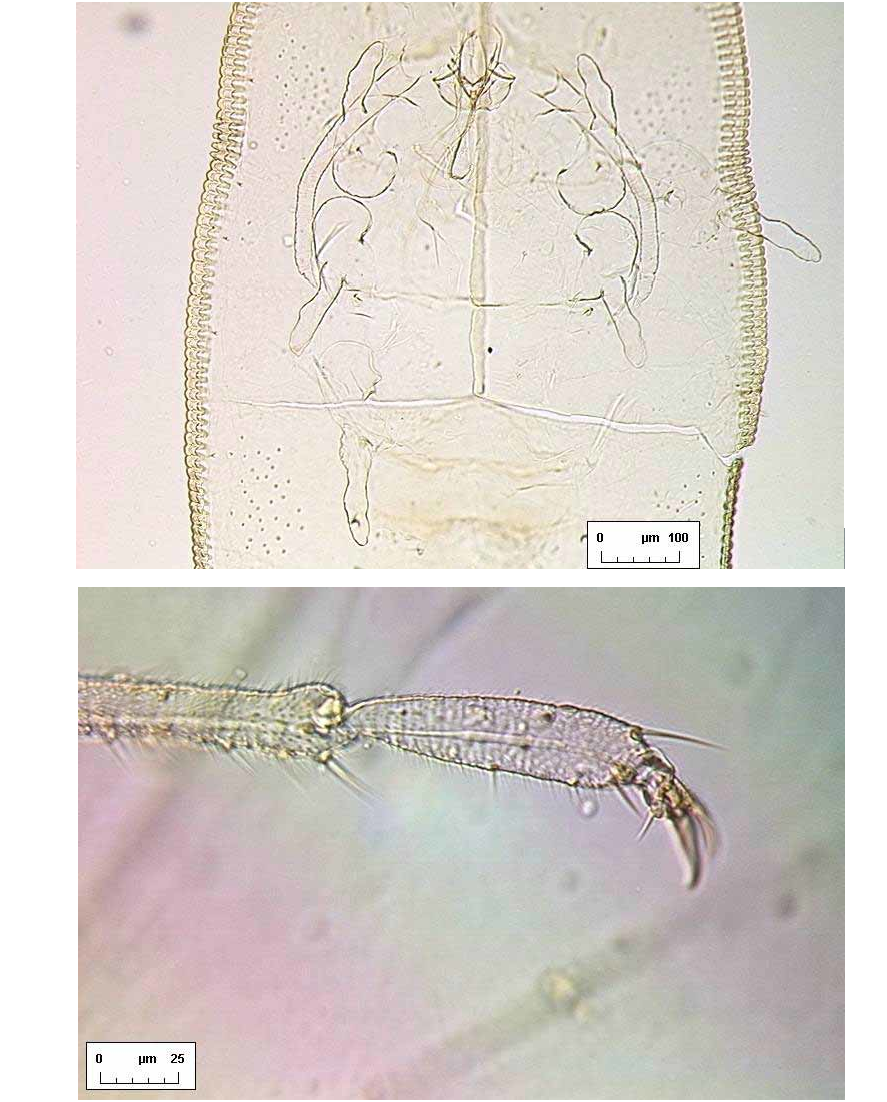
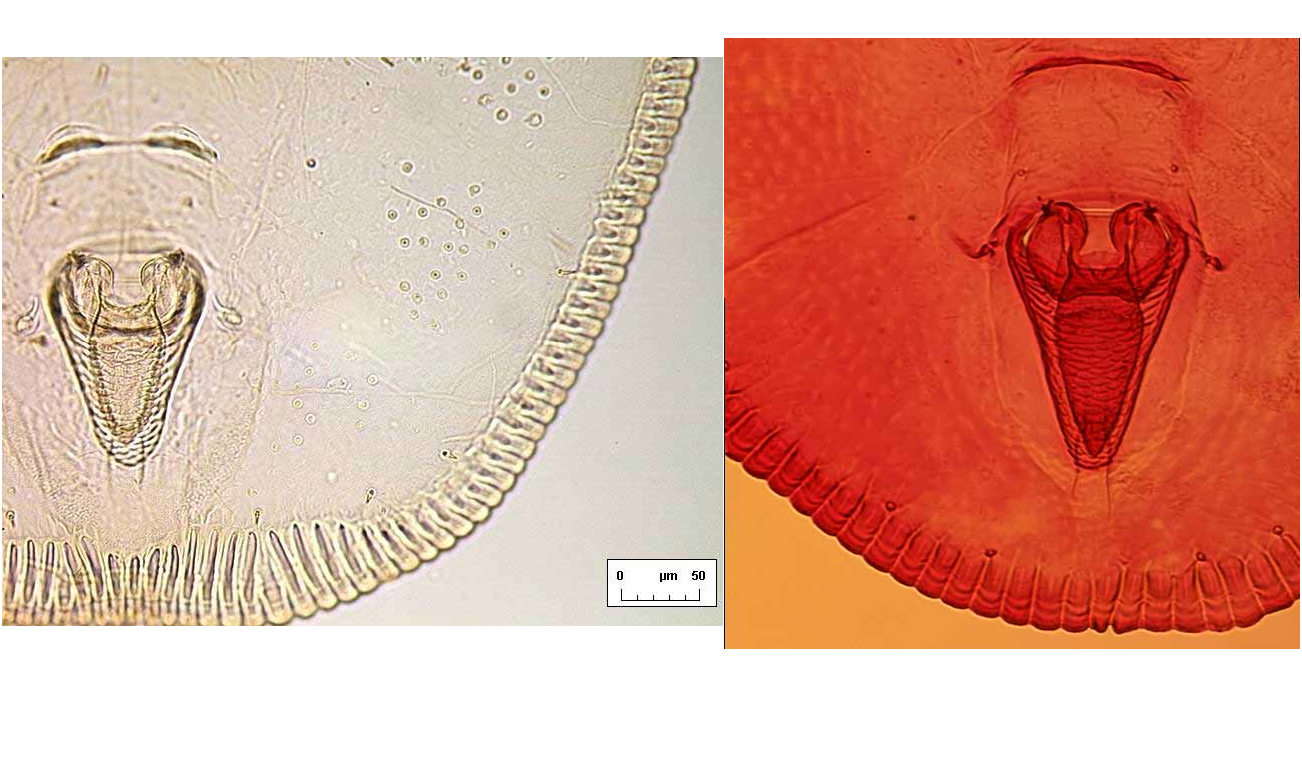
A B Dorsum. Longitudinal moulting suture reaches puparial margin; transverse moulting sutures almost straight, directed very slightly posterolaterad and reaching almost as far as the submarginal setae. About one marginal toothwidth inside puparial margin is a row of glandular crenulations closely resembling a second rank of marginal teeth, and exactly in register with them ( Fig. 6 View FIGURE 6 B); glandbase folds extend about one toothlength mesad, except at the puparial apices where the folds become much longer, those at the posterior end of the puparium extending about halfway to apex of vasiform orifice ( Fig. 7 View FIGURE 7 A). Dorsal disc almost smooth, but abdominal segments medially with transverse rows of tiny spinules which are most distinct posteriorly from segment IV. Meso / metathoracic and abdominal segmentation clear submedially ( Fig. 2 View FIGURE 2. S ), more subtly marked into subdorsum, abdominal segments all similar in length medially. Submedian abdominal depressions small, rounded, sometimes differentially staining. Vasiform orifice ( Fig. 3 View FIGURE 3 A) rounded triangular, about 1.5 times longer than wide, straightsided, its floor distinctly imbricate ( Fig. 7 View FIGURE 7 A), the orifice inset from posterior puparial margin by less than its own length; operculum ( Fig. 3 View FIGURE 3 B) transversely ovoid, its posterior margin shallowly “M”shaped and with a pair of setae; lingula head finely spinulose, straightsided, almost filling apical part of vasiform orifice, with two stout apical setae that extend beyond boundary of vasiform orifice, and two additional minute subapical setae that are difficult to distinguish from surface spinules but are usually visible with careful observation ( Fig. 3 View FIGURE 3 A). Caudal furrow absent, but vasiform orifice posterolaterally surrounded by a shallow “U”shaped ridge which abuts the posterior glandbase folds ( Fig. 7 View FIGURE 7 A).
Chaetotaxy. Anterior and posterior marginal setae present, spinelike, each placed on the flat surface of a marginal tooth, anteriors close to 4th cephalic pair of submarginal setae and posteriors between 4th and 5th posteriormost pairs of submarginal setae. Outer submargin normally with a row of 17 pairs of short, lanceolate spinelike setae ( Fig. 4 View FIGURE 4 A); single similar submedian pairs of cephalic, pro, meso and metathoracic and eighth abdominal setae also present, the eighth abdominals placed anterior to vasiform orifice, half way to abdominal pockets.
Pores. Compound pores entirely absent. A loose cluster of small dark pores ( Fig. 4 View FIGURE 4 B) occupies subdorsal sector of each of abdominal segments III–VIII ( Fig. 2 View FIGURE 2. S ), with a similar cluster on prothorax, and a with a few lining the submargin of the remaining cephalothoracic segments; some of the pores are circular, but others have a more complex and irregular outline. No other pores are apparent.
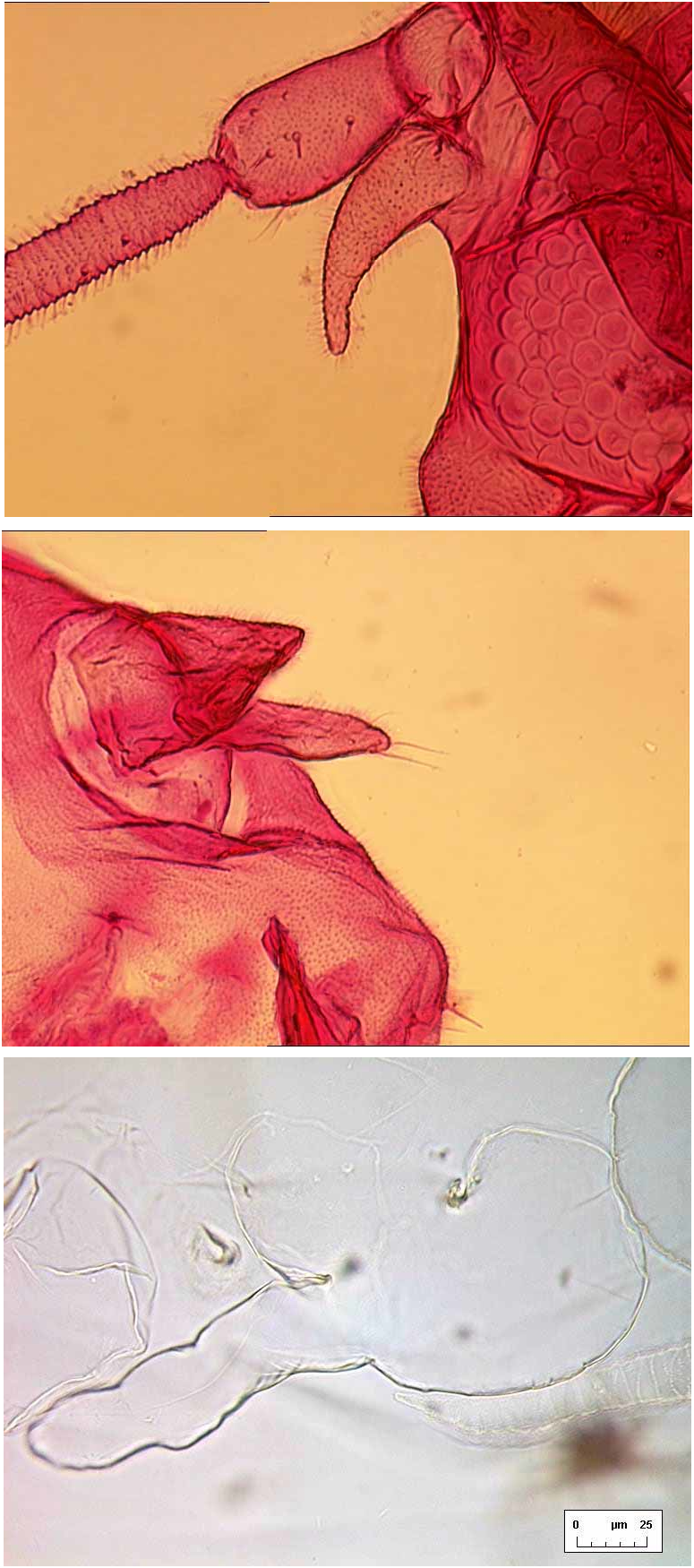
Ven te r. Cuticle delicate, smooth. Ventral abdominal setae fine, underlying vasiform orifice and operculum. Legs of highly characteristic appearance ( Fig. 10 View FIGURE 10 C), twosegmented and unusually elongate, the distal segments sinuatesided, directed anteriorly (fore legs) or posteriorly (middle and hind legs); legs each without an apical claw or pad, but sometimes a minute apical seta visible, and distal segments of middle and hind legs sometimes each with a tiny seta present on apical onethird part. Antennae placed anteromesad of fore legs, evenly curved back as far as middle leg articulations (females, Fig. 5 View FIGURE 5 A) or bases of hind legs (males). Caudal tracheal fold faintly marked by minute spinules; thoracic tracheal folds unmarked. A pair of tiny submedian setae is frequently visible underlying the tergite of abdominal segment II, often appearing dorsal but actually situated ventrally [this setal pair is more clearly visible, and unambiguously ventral, in puparia of S. vinsoni Takahashi in BMNH].
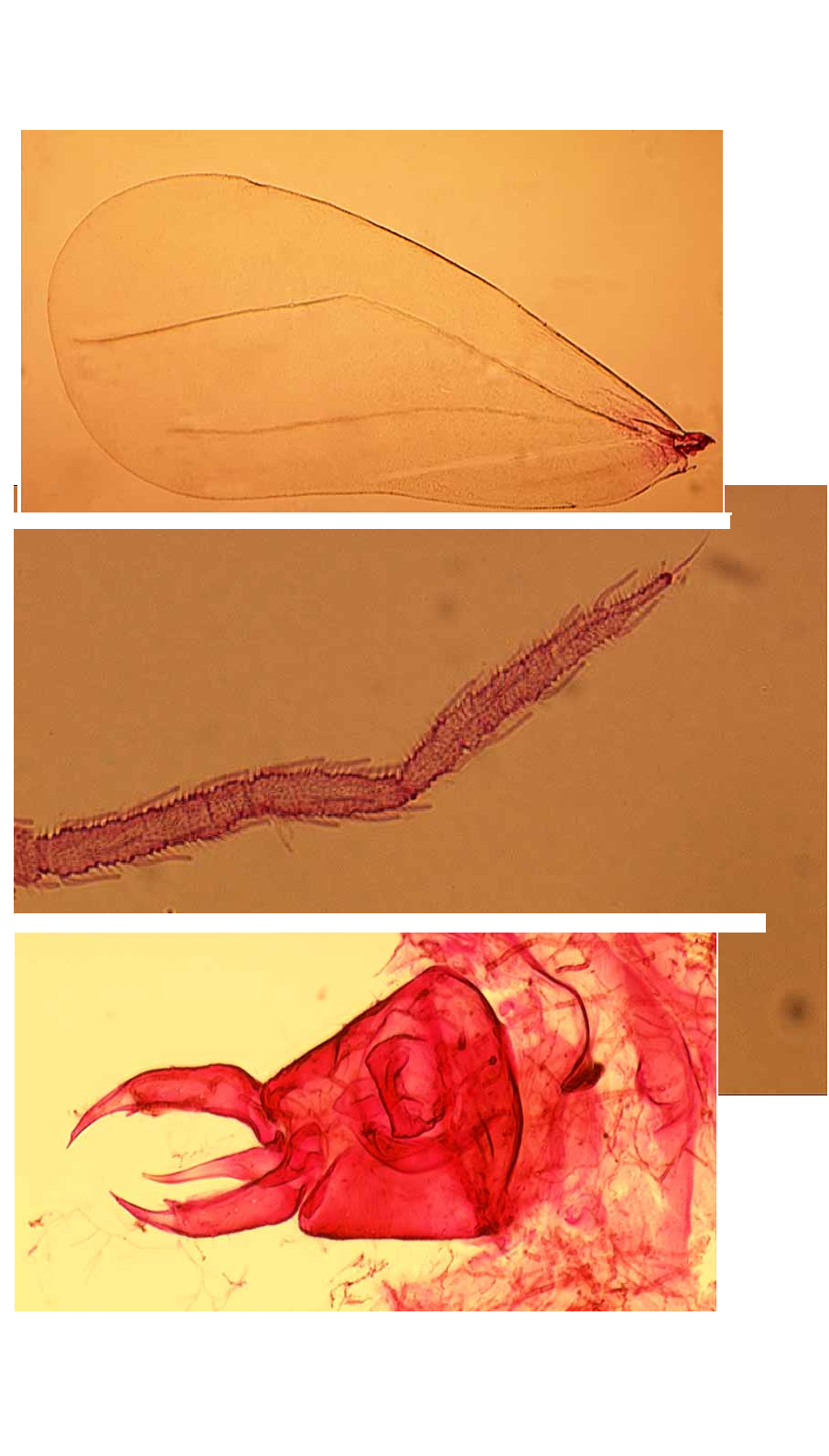
ADULT FEMALE ( Figs 5 View FIGURE 5 B, 6A, 8A, 9A, 10A, 10B). Body about 2.0 mm long (n=4). Fore wing ( Fig. 9 View FIGURE 9 A) elongate ovoid, vein R–Rs abruptly angled at halflength, with a very faintly indicated branch R1 sometimes indicated (by differential staining only), arising at the angle and directed towards the costal margin; pterostigma not indicated but vein Sc skirting costal margin basally; veins M, Cu and A also present (following the vein nomenclature of Solomon (1935) and Gill (1990)). Hind wing with R more convincingly developed into branches R1 and Rs than on fore wing, and also M distinct. Antennae 7 segmented, 0.65–0.70 mm long, the basal 3 segments thicker than the remainder, segment VII with a fine apical spine. Between antennal base and eye is an unusual curved, fingerlike, spinulose process ( Fig. 10 View FIGURE 10 A), of similar length to second antennal segment. Ultimate rostral segment very short, about as broad as long (6A). Each pair of tarsal claws with a ventrallydirected needlelike paronychium between them, visible in lateral aspect (5B).
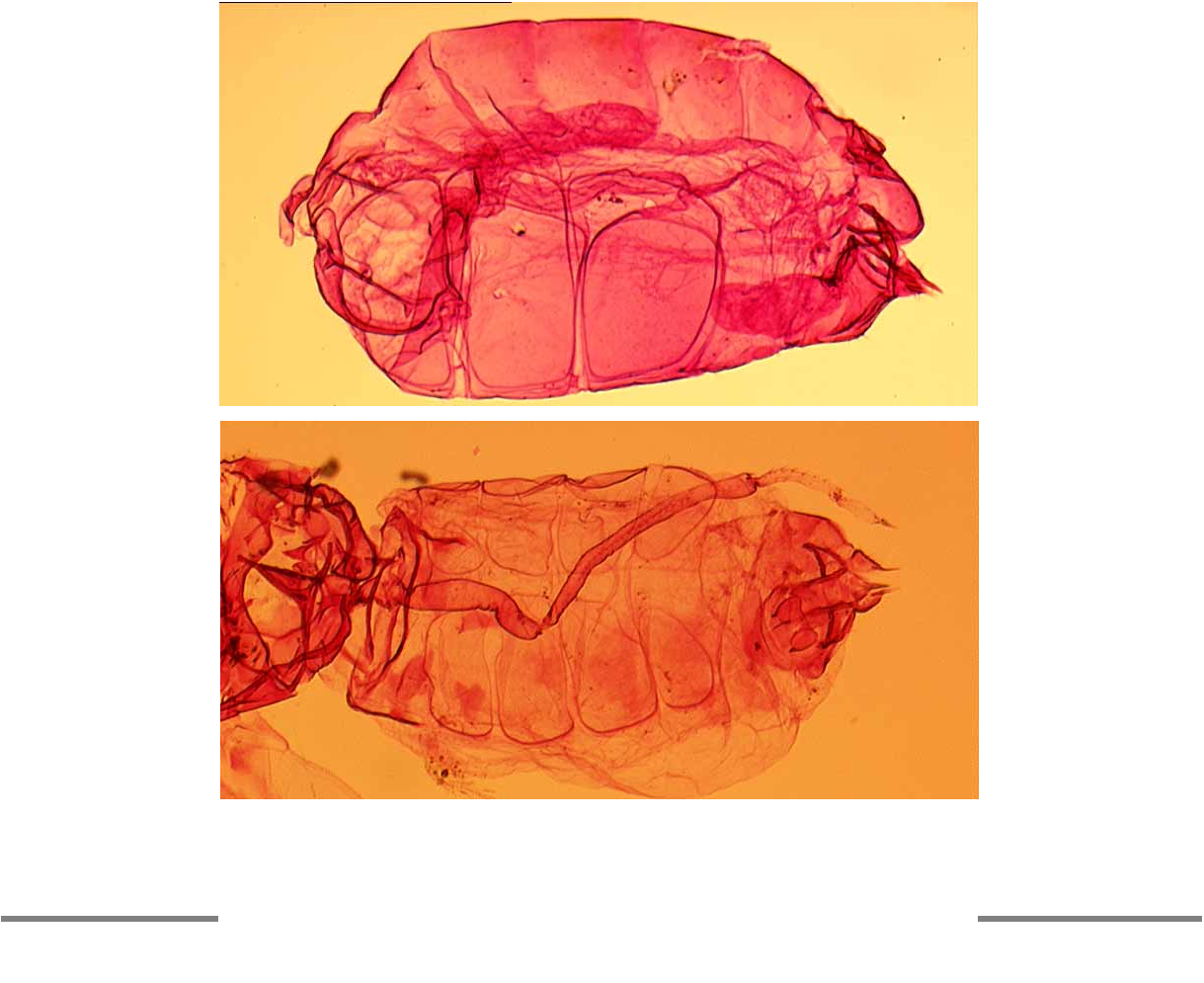
Abdomen with 3 pairs of large ventrolateral wax plates ( Fig. 8 View FIGURE 8 A), the fine individual facets resolvable at magnification factors of X250 and over. Vasiform orifice subcircular, almost fully occupied by trapezoidal operculum which has its pair of posterior marginal setae visible; lingula extends beyond boundary of vasiform orifice, with 4 small setae visible ( Fig. 10 View FIGURE 10 B).
ADULT MALE ( Figs 9 View FIGURE 9 B, 9C). As for female except as follows. Body 1.55–1.70 mm long (n=4). Claspers ( 0.15 mm) and aedeagus ( 0.12–0.13 mm) much thickened basally, narrowly acute apically ( Fig. 9 View FIGURE 9 C). All specimens heavily obscured by fungus, but can be seen to have 4 pairs of ventrolateral wax plates, of similar structure to those in the female, and this is further supported by a preemergence male inside one of the mounted puparia. There is a median ventral tubercle immediately posterior to the abdominal wax plates, bearing a few setae. Only one complete antenna is present, 7segmented, 0.60 mm long, the flagellum ( Fig. 9 View FIGURE 9 B) densely provided with blunt setalike sensoria (termed uncinulae by BinkMoenen, 1983).
MATERIAL EXAMINED. HOLOTYPE, puparium, MAYOTTE ISLAND, Dani, on Cocos nucifera (Palmae) , 09.vii.2003 (Laurence Ollivier 0300462) ( MNHN). PARATYPES, 33 puparia, 2 adult males, same data as holotype ( BMNH, ENSAM, MNHN); 35 puparia, 3 thirdinstar larvae, 1 secondinstar larva, 5 adult females, 3 adult males, same host and locality, 10.vii.2003 (Ollivier 0300475); 6 puparia, 3 adult females, 7 puparia, on Cocos nucifera, Coconi, 2003 (Ollivier) ( BMNH, ENSAM, MNHN, USNM); 15 puparia, 2 thirdinstar larvae, TANZANIA [“ Tanganyika Territory ”], Bagamoyo, on Cocos nucifera , 18.vi.1936 (D. Vezey Fitzgerald 5054) ( BMNH); 2 puparia, KENYA, Matuga, on Cocos nucifera , 08.ii.1982 (Kibata; Commonwealth Institute of Entomology A.13866) ( BMNH). OTHER MATERIAL, numerous puparia from Mayotte, dry on leaves, same data as above ( BMNH, ENSAM).
ETYMOLOGY. The specific epithet is the French word papillote , a special Christmas twistwrapped candy, which puparia of this new species resemble by virtue of the most unusual periferal wax secretions. The word is used as noun in aposition.
DISCUSSION. The puparia of S. papillote differ from those of S. vinsoni in several respects. The submarginal glands in S. vinsoni do not so much resemble a second rank of marginal teeth; the toothbase folds are not longer at the anterior and posterior ends of the puparium in vinsoni ( Fig. 7 View FIGURE 7 B); the posterior opercular margin of vinsoni is not “M”shaped as in papillote ; vinsoni has distinct clusters of papillae skirting the inner boundary of the marginal toothbase folds, and these are absent in papillote ; the dorsal setae in vinsoni are not lanceolate as they are in papillote ; the subdorsum in vinsoni is not punctuated by groups of dark simple pores as in puparia of papillote . The puparia of both species suggest inclusion of Stenaleyrodes in the Aleurodicinae, with four lingular setae visible under high magnification (X250 or X400).
The only adult specimens of S. vinsoni available for study are preemergence individuals dissected from puparia from New Caledonia, and these have four lingular setae, as also seen in papillote . However, these preemergence adults clearly display one major difference from the females of papillote , in that both sexes of vinsoni have four pairs of ventrolateral wax plates, compared with only three pairs in females ( Fig. 8 View FIGURE 8 A) and four pairs in males of papillote . Fig. 8 View FIGURE 8 B clearly shows the four paired wax plates in a preemergence female of S. vinsoni . Although Gill (1990) stated that adults of the Aleurodicinae possess four pairs of abdominal wax plates in females, and three pairs in males, this character is not reliable. Branching of vein R into R1 and Rs in whiteflies is sometimes a function of larger physical size. In Stenaleyrodes this branching is only sometimes faintly indicated by differential staining, but the presence of other additional veins strongly suggests inclusion in the Aleurodicinae. The fore wings of S. papillote , and Dumbleton’s rather over simplified illustration of the fore wing of D. elongatus , bear a great resemblance to those of an Australian genus, Synaleurodicus Solomon (1935) , differing principally in the more poorly defined branch R 1 in S. papillote . However, as discussed in detail by Martin (1999), Synaleurodicus is accommodated in the Aleurodicinae with considerable caution, being even more atypical than Stenaleyrodes .
The nature of the puparial secretions of S. papillote ( Fig. 1 View FIGURE 1 ) deserves comment. The fringe comprises coalesced filaments which are secreted by the contiguous submarginal glands, in a manner that is common amongst many other whitefly species. What is highly unusual in S. papillote is the way that almost all these filaments are perpendicular to the puparial axis, despite the curved margin: in virtually all other fringed whitefly puparia, the fringe forms an even annulus surrounding the body, rather than the remarkable longitudinal apical “brushes” and lateral “skirts” of S. papillote .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |