
Knodus rufford, Deprá & Ota & Vitorino Júnior & Ferreira, 2021
|
publication ID |
https://doi.org/10.1590/1982-0224-2020-0106 |
|
publication LSID |
lsid:zoobank.org:pub:C4F52922-98BE-485C-94F7-03D158B4EDEB |
|
DOI |
https://doi.org/10.5281/zenodo.10992426 |
|
persistent identifier |
https://treatment.plazi.org/id/BC88B912-CF9A-47EA-ABFF-746043DCD06F |
|
taxon LSID |
lsid:zoobank.org:act:BC88B912-CF9A-47EA-ABFF-746043DCD06F |
|
treatment provided by |
Felipe |
|
scientific name |
Knodus rufford |
| status |
sp. nov. |
Knodus rufford , new species
urn:lsid:zoobank.org:act:
( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ; Tabs. 1–2 View TABLE 1 View TABLE 2 )
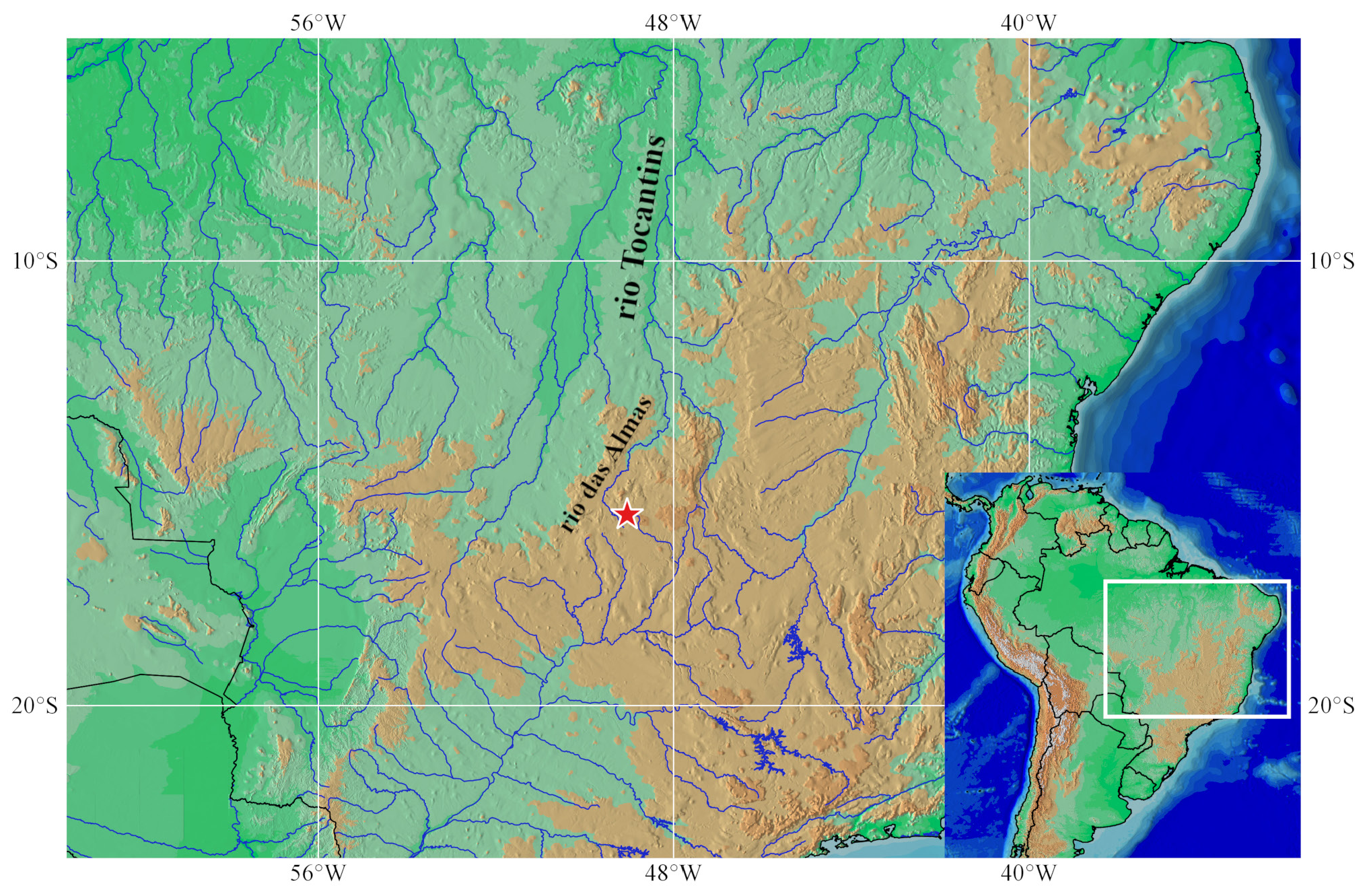
Holotype. NUP 22661, 35.4 mm SL, Brazil, Goiás State, Municipality of Pirenópolis, ribeirão Dois Irmãos, tributary to the rio do Peixe , tributary to the rio das Almas, tributary to the rio Maranhão, tributary to the rio Tocantins, 15°42’50”S 49°2’39”W, elevation 713 m, 12 Dec 2016, G. C. Deprá & O. Vitorino Júnior. GoogleMaps
Paratypes. All from Brazil, State of Goiás, Goiás State, Municipality of Pirenópolis , ribeirão Dois Irmãos , tributary to the rio do Peixe , tributary to the rio das Almas, tributary to the rio Maranhão, tributary to the rio Tocantins. CPUFMT 6834 , 1 c&s, 33.1 mm SL, collected with holotype. GoogleMaps CPUFMT 6835 , 2 , 25.2–31.7 mm SL, 15°42’57”S 49°52’37”W, elevation 715 m, 12 Dec 2016, G. C. Deprá & O. Vitorino Júnior. GoogleMaps MCP 54464 View Materials , 1 View Materials , 31.8 mm SL, 15°42’50”S 49°2’54”W, elevation 702 m, 12 Dec 2016, G. C. Deprá & O. Vitorino Júnior. NUP 22660 , 28.2 – 36.8 , collected with holotype. GoogleMaps NUP 22669 , 4 , 28.1–39.2 mm SL, type locality, 24 Apr 2017, G. C. Deprá & O. Vitorino Júnior GoogleMaps .
Non-type. CPUFMT 6833, 1 (fixed in 90% ethanol), 34.4 mm SL, Brazil, Goiás State, Municipality of Pirenópolis, ribeirão das Araras, tributary to the ribeirão Dois Irmãos, tributary to the rio do Peixe , tributary to the rio das Almas, tributary to the rio Maranhão, tributary to the rio Tocantins, 15°42’15”S 49°2’12”W, elevation 722 m, 12 Dec 2016, G. C. Deprá & O. Vitorino Júnior GoogleMaps .
Diagnosis. Knodus rufford differs from all other congeners bearing caudal-fin scales, except K. figueiredoi , and from Bryconacidnus hypopterus , by having up to three cusps in the premaxillary inner row teeth and dentary teeth (rarely a fourth or fifth cusp present in one or two premaxillary teeth). Knodus rufford differs from K. figueiredoi by having 4–4½ scales between dorsal-fin origin and lateral line ( vs. 5), 11–12 circumpeduncular scale rows ( vs. 13–14), and i,6,i pelvic-fin rays ( two specimens with i,5,i; vs. always i,5,i). Additionally, the number of scale rows between dorsal-fin origin and lateral line distinguishes Knodus rufford from all congeners bearing caudal-fin scales, except K. cupariensis , K. meridae , K. nuptialis , K. orteguasae and smaller specimens of the syntopic K. aff. breviceps and K. obolus (new species, described below), and from Bryconacidnus hypopterus . The number of circumpeduncular scale rows distinguishes K. rufford from all congeners bearing caudal-fin scales, except K. angustus , K. borki , K. cupariensis , K. delta , K. longus , K. nuptialis , and K. pasco , and from B. hypopterus . The completely pored lateral line distinguishes K. rufford from K. borki (incomplete, 6–14 perforated scales) and K. delta (8–12). The absence of distinct marks from all fins distinguishes K. rufford from K. dorsomaculatus (a black band present on proximal third of dorsal fin), K. cupariensis and K. geryi (a black spot on the base of each caudal-fin lobe), K. pasco (a horizontal black band present on the middle of dorsal fin) and K. savannensis (a distinct black band running along middle caudal fin rays and extending through distal half of ventral caudal-fin lobe). The body depth (28.6–34.7% SL) distinguishes K. rufford from K. angustus (20.0–25.2), K. longus (22.5–23.7), K. megalops (34.2–41.6) and K. obolus (36.6– 40.1). The presence of 1–4 maxillary teeth distinguishes K. rufford from K. pasco (5–6). The presence of 2½–3 scales between lateral line and pelvic-fin origin distinguishes K. rufford from K. chapadae (3½–4) and K. shinahota (5). The presence of normally developed skin folds on the base of the first few branched anal-fin rays ( vs. highly developed) and the absence of a second humeral blotch ( vs. presence) distinguishes K. rufford from K. tiquiensis . The absence of breeding tubercles distinguishes K. rufford from K. nuptialis . Other characters, although shared with several other congeners, help distinguishing Knodus rufford . The number of branched anal-fin rays (15–17) distinguishes K. rufford from all congeners bearing caudal-fin scales, except K. angustus , K. breviceps , K. chapadae , K. figueiredoi , K. geryi , K. heteresthes , K. longus , K. meridae (more frequently 13 or 14), K. mizquae , K. orteguasae , and K. tiquiensis , and from B. hypopterus . The number of scales in the longitudinal series (34–36, all of which are perforated in K. rufford ) distinguishes K. rufford from all congeners bearing caudal-fin scales, except K. borki (lateral line incomplete), K. cupariensis , K. delta (lateral line incomplete), K. figueiredoi , K. gamma , K. heteresthes , K. megalops , K. meridae , K. nuptialis , K. orteguasae , K. savannensis , K. shinahota and K. victoriae .
Description. Morphometric data in Tab. 1 View TABLE 1 . Body compressed, greatest body depth anterior to vertical through dorsal-fin origin. Dorsal profile of body convex from upper lip to vertical through nostril; slightly convex from this point to base of supraoccipital bone; slightly concave from this point to distal tip of supraoccipital process. Ascending, slightly convex from tip of supraoccipital process to dorsal-fin origin; descending, straight along dorsal-fin base; continuing almost straight to adipose-fin origin and along caudal peduncle. Ventral profile of body convex from lower lip to anal-fin origin; straight along anal-fin base; and concave to almost straight along caudal peduncle.
Jaws slightly retrognathous, mouth terminal. Premaxillary teeth in two rows. Outer row with 3(3), 4*(6), or 5(2) conical to tricuspid teeth. Inner row with 4*(11) teeth, all of them tricuspid in eight specimens; in the holotype, the third tooth (from symphysis) is pentacuspid; in one specimen ( 28.8 mm SL), the symphyseal tooth is tetracuspid; and in one specimen ( 28.2 mm SL), the second and the third teeth are tetracuspid. Posterior margin of maxilla posterior to vertical through anterior margin of orbit. Maxilla with 1(1), 2(2), 3*(7), or 4(1) tricuspid teeth. Dentary with 3(1), or 4*(10) tricuspid teeth, gradually decreasing in size, followed by up to seven conical and abruptly smaller teeth. Central cusp longer than lateral ones.
Scales cycloid, with few radii (3–5); circuli markedly anterior and marginally (dorsally and ventrally). Lateral line completely pored with 34(1), 35*(7), or 36(2) perforated scales. Scale rows between dorsal-fin origin and lateral line 4(6), or 4½*(4). Scale rows between lateral line and pelvic-fin insertion 2½(8), or 3*(3). Predorsal series with irregular pattern, uniserial with 9(1), 10(2) scales, or bisserial on anterior portion and uniserial on posterior portion, 6*(4), 7(3), 8(1), 10(3) scales in midline. Single row of up to 15 scales covering base of anal-fin rays. Circumpeduncular scales 11*(4) or 12(4). A single series of 13(2), 14(1) scales covering the entire anal-fin base.
Dorsal-fin rays ii,6,ii (1; penultimate element in process of bifurcation) or ii,7,i*(10). Adipose fin present. Pectoral-fin rays i,9,i(2), i,9,ii(1), or i,10,i*(8), distal tip of rays not reaching pelvic-fin origin. Pelvic-fin rays i,6(1), or i,6,i*(10), distal tip reaching anal-fin origin. Anal-fin rays iii,15(3), iii,16*(7), or iii,17(1), as seen in entire specimens (but see Tab. 2 View TABLE 2 for c&s specimen). Caudal fin bifurcate, lobes approximately of same size, i,9/8,i rays.
First gill arch with 4(2), or 5*(9) external rakers on upper limb, 1(11) on intermediate cartilage, and 8(8), 9(2), or 10*(1) on lower limb, as can be seen in entire specimens. For a more detailed gill-raker count of the c&s specimen, see Tab. 2 View TABLE 2 . Branchiostegal rays 4(11).
Osteology. General appearance of c&s specimen shown in Fig. 4 View FIGURE 4 . Meristic osteological characters are summarised in Tab. 2 View TABLE 2 . Rhinosphenoid ossified ( Fig. 4 View FIGURE 4 ); epiphyseal branch of the laterosensory canal present. Contralateral frontals contacting along about 90% of distance between mesethmoid and epiphyseal bar, with left frontal bearing expansion over right one. Anterior fontanelle length less than 10% posterior fontanelle length. Posteromedial corner of parietal rounded. Parasphenoid bar slightly arched. Anteroventral process of mesethmoid well developed, nested between contralateral premaxillae. Pharyngobranchial 2 toothless, much shorter than pharyngobranchial 3. Ossifications dorsal to cartilage between basibranchials 2–3 absent. Ceratohyal with deep notch at the articulation with branchiostegal rays 2–3.
Paratypes
Holotype N Range Mean SD 36.8 10 25.2–39.2 31.4 - Percentages of standard length Body depth 34.2 10 28.6–34.7 31.6 2.2 Predorsal distance 54.9 10 52.3–55.7 54.3 1.3 Prepectoral distance 23.9 10 24.3–26.6 25.1 0.7 Prepelvic distance 46.7 10 46.8–49.7 47.9 1.0 Preanal distance 63.3 10 60.1–64.0 62.3 1.3 Caudal-peduncle depth 12.0 10 10.2–13.3 11.4 1.0 Caudal-peduncle length 13.6 10 12.6–14.1 13.4 0.5 Dorsal-fin base length 11.7 10 11.0–13.5 12.2 0.8 Dorsal-fin length 24.5 10 22.2–26.2 24.4 1.3 Pectoral-fin length 21.5 10 20.9–22.8 21.9 0.5 Pelvic-fin length 15.2 10 14.3–16.7 15.1 0.7 Anal-fin base length 26.6 10 24.5–28.0 26.6 1.4 Anal-fin length 17.9 10 16.1–18.8 17.3 1.0 Orbit to dorsal-fin origin 42.1 10 38.8–44.1 41.6 1.4 Dorsal-fin origin to caudal peduncle 50.0 10 45.9–51.8 49.4 1.8 Head depth 25.0 10 23.4–25.5 24.6 0.6 Head length 24.7 10 24.8–26.8 25.6 0.7 Percentages of head length Orbital diameter 37.4 10 36.2–39.8 38.2 1.2 Snout length 28.6 10 24.2–26.7 26.0 0.7 Interorbital width 37.4 10 34.1–39.0 37.3 1.4 Upper-jaw length 37.4 10 36.2–38.4 37.4 0.6
Colour in alcohol. Background colour beige to pale yellow. Dorsal region of head and mid-dorsal region of body with dense concentration of melanophores. Iris with scattered melanophores. Region of infraorbital and opercular series beige, with few scattered melanophores, more concentrated on superior half. Region of opercular series retaining guanine pigmentation. Maxilla and gular region with few scattered melanophores, slightly more concentrated on superior half. Flank with melanophores scattered and equally distributed, except on anteroventral half, and on ventral portion of caudal peduncle. Scales on superior third of anterior region of flank with dense concentration of melanophores on posterior margin, conferring reticulate pattern. One dark-brown humeral blotch, conspicuous, shaped as square or vertically elongate rectangle, occasionally with margins somewhat rounded; with equally distributed melanophores, extending two longitudinal scale rows above lateral line and not surpassing it below. Brownish longitudinal midlateral stripe, from vertical through halfway humeral blotch and dorsal-fin origin, to median caudal-fin rays. Fins almost hyaline, with few melanophores on interradial membranes; dark-brown stripe along anal-fin base. Abdominal region, from pectoral-fin origin to pelvic-fin end, without melanophores.
Sexual dimorphism and ontogeny. Two specimens collected in December and April ( 28.1–31.7 mm SL; presumably males) with contact organs (hooks) on anal and pelvic fins and gill-derived gland. In both specimens, hooks present on distal half of largest unbranched and first three branched anal-fin rays (4–6 small, paired hooks per ray, one per segment, on posterolateral side of each ray, most proximal situated immediately proximal to first branching point and more distal ones located only on posterior branch, if on branched ray); and in lateralmost unbranched and in following four branched pelvic-fin rays (7–12 hooks per ray, one ray per segment, on ventromedial side of each ray, most proximal situated well proximal to first branching point and more distal ones located only on medial branch, if on branched ray). Gill-derived gland formed by fusion of 6–8 most anteroventral external filaments of first gill arch, although former limits between them still obvious. No allometric nor allomeric changes detected, possibly
due to small available sample. Allochromatic changes include increase in contrast of colour patterns and overall increase in number of melanophores on body. Longitudinal midlateral stripe progressively more intense, deeper and with sharper limits. Also an increase in concentration of guanine and of melanophores dispersed around midlateral stripe, mainly in region between humeral mark and vertical through dorsal-fin origin. Humeral mark also develops sharper and more squarish limits (more diffuse and roundish in young). Countershading, which in young consists only in a few melanophores on distal margin of dorsalmost scales, much more developed in adults. In some adults, first few lateral-line scales with pores bordered by melanophores. No apparent sexual dichromatism.
Etymology. We name Knodus rufford after the Rufford Foundation, which provided the funds that allowed the expedition resulting in the discovery of the two new species described herein (as well as Moenkhausia goya Deprá, Azevedo-Santos, Vitorino Júnior , Dagosta, Marinho & Benine). A noun in apposition not latinised.
Geographical distribution. Knodus rufford is only know from the ribeirão Dois Irmãos basin, in the rio das Almas basin, upper rio Tocantins, Municipality of Pirenópolis, Goiás State, Brazil ( Fig. 5 View FIGURE 5 ).
Ecological notes. Knodus rufford occurs in all sorts of aquatic environments within the high-altitude (about 700 m) streams of the rio das Almas basin, despite there seems to be some preference for pool habitats where the water flow is less intense and most of the individuals were sampled. Stomach content analyses indicate omnivorous behaviour once both species consumed aquatic and terrestrial invertebrates, plant material, organic and inorganic debris. Disturbances such as habitat degradation and rapid increase of water flow during the rainy season were often observed on sampling sites with occurrence of the species, indicating population-level responses that increase species tolerance to variability of habitat conditions, resources availability and other mortality factors. Therefore, K. rufford fits the classic opportunistic life-history model (Winemiller, 1989) with less specialized requirements in every dimension of its niche.
Conservation status. Knodus rufford is so far only known from the rio das Almas system, an upper rio Tocantins basin tributary. Inside this watershed the species is widely distributed and was recorded at relatively degraded habitats mainly by pasture and mining activities. As no specific threats were detected, K. rufford can be tentatively categorized as Least Concern ( LC) following the International Union for Conservation of Nature ( IUCN) categories and criteria ( IUCN Standards and Petitions Subcommittee, 2017).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |