
Uromys caudimaculatus (Krefft, 1867)
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.46.1994.12 |
|
DOI |
https://doi.org/10.5281/zenodo.4658403 |
|
persistent identifier |
https://treatment.plazi.org/id/DD3387EC-FFDF-6654-72DD-3DA6F997FC5D |
|
treatment provided by |
Felipe |
|
scientific name |
Uromys caudimaculatus (Krefft, 1867) |
| status |
|
Uromys caudimaculatus (Krefft, 1867) View in CoL
Type material. SYNTYPE, AM S1848 , skull only (the body apparently being lost). Cape York .
Revised diagnosis. Uromys caudimaculatus differs from U. anak and U. neobritannicus in possessing a posterior palatal spine, an only rudimentary postorbital process; narrow, tapering posterior nasals which terminate anterior to the posterior ends of the lachrymals; in having the hindfoot not usually longer than 22% of the head-body length and ear length more than 43% of condylobasal length; the incisive foramina are more than 20% of the palatal length, while the toothrow is less than 20% of condylobasal length. The colour is a medium brown-grey, the feet are white, the tail scales arranged in clear rings and longer than broad, with a white tail tip. It differs from U. hadrourus in its larger size, less inflated braincase, and larger postorbital process and anterolateral bullar spurs, shorter ears and longer white tail tip, greater frontal convexity, narrower feet and less posteriorly extensive nasals. It differs from U. emmae n.sp. in its more elongate rostrum, narrower and longer hindfoot, relatively smaller teeth, longer ears, more inflated frontals, and in having the distalmost portion of the tail white.
Notes. We have considered whether there might not be two or even three distinct species among what was hitherto called U. caudimaculatus . As far as the mainland (Australian and New Guinean) forms are concerned, this might be possible. The work of Donnellan (1989), based upon chromosomes, suggests that these populations may represent distinct species, since a possible rearrangement (tandem fusions) in the Australian population should provide barriers to gene flow. C-banding in the New Guinean population, however, is necessary before tandem fusion can be identified with certainty. A morphological study also shows that a distinct form inhabits Australia and southern New Guinea as far east as Kaimare in the Gulf of Papua, a second homogeneous taxon is found throughout the rest of the eastern half of New Guinea, and a third form is found in the western part of the island. Unfortunately, the characters that support this division are not entirely consistent, and when the insular populations are taken into consideration they form such a complex mosaic pattern that it is impossible to allocate the insular forms satisfactorily to one or other of the mainland species. Thus, while recognising the striking differences between the three mainland forms, and the lack of intermediates in some regions on the mainland, we have declined to recognise them as full species, but instead regard them as subspecies. Study of further material will be necessary to confirm or refute this hypothesis.
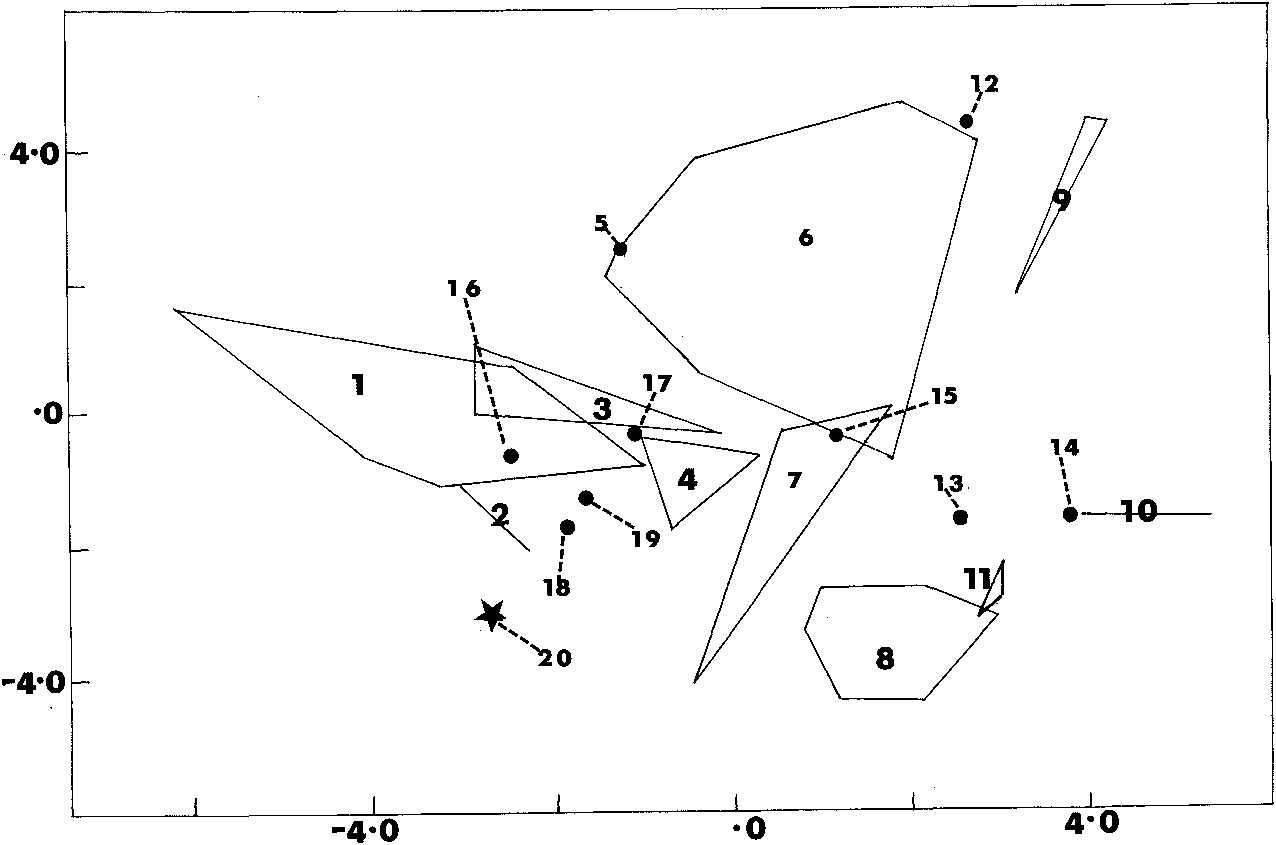
The Discriminant Function analysis ( Fig. 4 View Fig ) tends to separate the three major mainland groups, but not cleanly. Clustering with the Australian / south New Guinea sample are specimens from Yapsiei and Yapen Island , and an Am Islands specimen is in the slight overlap zone between the Australian / south New Guinean and the main Papua New Guinean samples. The Irian Jayan samples are close to one another, but a specimen from Kaimare (which in most respects apart from its metrical characters resembles the Australian form) falls within the Irian Jayan cluster. The only taxon recognised here as a full species that does fall cleanly outside any major cluster is the holotype of U. emmae n.sp .
Within Papua New Guinea there is a series of overlapping clusters: Mount Hagen area/south-east PapuaIMount SisalDobodura. Specimens from Telefomin and Mount Karimui fall on the edge of the Mount Hagen cluster; one from the Torricelli Mountains falls, curiously, with the Mount Sisa specimens. One Mount Elimbari specimen falls outside any of these. Within Irian Jaya there is again a gradient: Alkmaar district! Setakwa/Mimika RiverlUtakwa, with a specimen from the Weyland Range falling, oddly, in the Papua New Guinea cluster.
| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.