
Pelobatidae Bonaparte
|
publication ID |
https://doi.org/10.11646/zootaxa.3661.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:448C4455-5A22-4C99-AA04-6FAF6DAFB879 |
|
persistent identifier |
https://treatment.plazi.org/id/B64F87EA-2079-FFE5-FF20-2DA459EDBFD3 |
|
treatment provided by |
Felipe |
|
scientific name |
Pelobatidae Bonaparte |
| status |
|
Family Pelobatidae Bonaparte View in CoL
Moroccan Spadefoot Toad, Pelobates varaldii Pasteur and Bons 1959
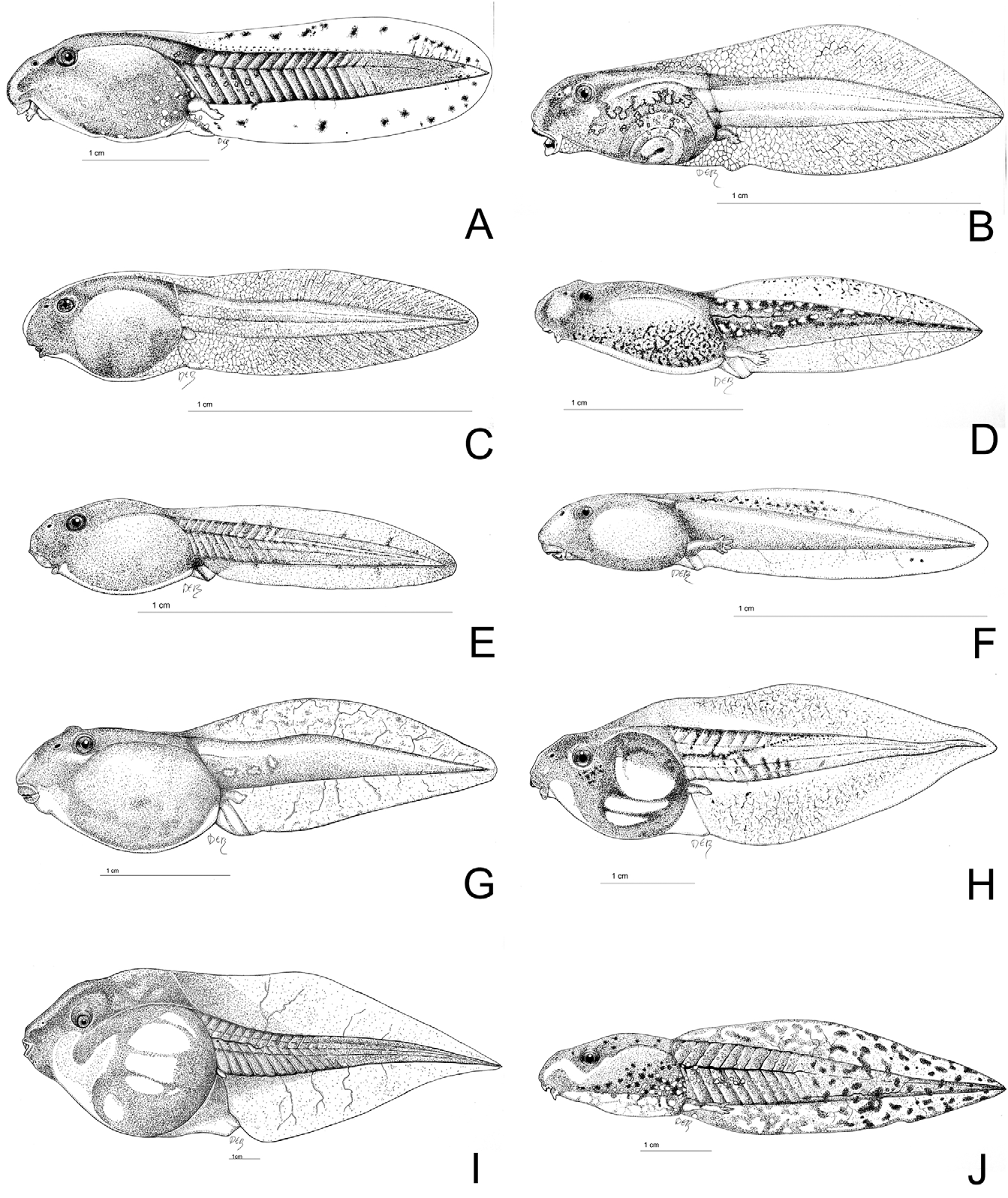
Figs. 15D–F View FIGURE 15 .
Background information. Information on P. varaldii remained scarce after the discovery of the species until Busack et al. (1985) provided data on the electrophoretic, immunological and morphological discrimination between P. varaldii and P. cultripes , which showed little morphological but considerable genetic differentiation. Recently, García-París et al. (2003) and Veith et al. (2006) found a well-supported sister taxa relationship between P. varaldii and P. cultripes , which dates back to the end of the Messinian Salinity Crisis, approximately 5.3 mya. Conclusively, both species likely diverged as a result of the collapse of the Gibraltar land bridge. Recent contributions by Crochet and Geniez (2003), Guzman et al. (2007), El Hamoumi et al. (2007), Guarino et al. (2011) and de Pous et al. (2012) have revealed details on the distribution, phenology and natural history of P. varaldii . Confusion regarding species-specific morphology however remained, as Pasteur and Bons (1959) described P. varaldii adults as well as larvae to be highly divergent from P. cultripes based on a large sample of individuals, while Busack (1985) considered adults of both species to be highly similar in external morphology. However, the P. varaldii individuals analysed by Busack (1985) were partially substituted with small P. cultripes due to the low sample size of the former. Despite the fact that substrate is known to influence at least the size of Pelobates spp. (i.e. individuals located on sandy substrates tend to remain smaller; Marangoni & Tejedo 2008; Marangoni et al. 2008), descriptive statistics indicate that P. varaldii has a smaller mean and maximum SVL length compared to P. cultripes as reported in the available literature ( Table 4). In addition to these results, we confirm the abundant presence of red dots (absent in P. cultripes ), most pronounced on the upper eyelids but often also present on the forelimbs, hindlimbs, dorsum (sometimes including flanks) and head ( Pasteur & Bons 1959, see Fig. 15D, F View FIGURE 15 ) in the majority of individuals ( Table 5). Furthermore, larval morphology significantly differs between these two sister species as elaborately described by Pasteur and Bons (1959). Altogether, we regard the degree of morphological (larval and adult) and genetic divergence sufficient to recognize P. varaldii and P. cultripes as different species.
Tadpole. Pasteur and Bons (1959) gave an elaborate description of the tadpole of P. varaldii as support for specific status of this taxon. A summary of their results is presented here. A drawing of the tadpole is displayed in Fig. 12I View FIGURE 12 . Fully developed tadpoles are characterized by their large size (up to 130 mm, occasionally larger), globular body and presence of black metatarsal tubercles. Eyes positioned laterally on head. Body length covers approximately 1.3 to 1.8 times body height and 1.6 times its width. Dorsum flat. Length of the tail ranges between 1.38 to 2.08 times body length. Tail length 1.8 to 2.4 times longer than deep. Dorsal and ventral tailfin highly extended, tapering to a point at the tip of the tail. Spiracle sinistral, pointing diagonally upwards. Oral disk oval shaped. Four or five tooth rows on the upper and five on the lower labium. Upper supralabial row highly reduced, large median gap in other supralabial rows, interspersed by mouth. Median gap in all infralabial rows except the lowest row. Labial rows with median gaps always asymmetrically divided. Colour uniform grey, the dorsum being somewhat darker in comparison with the rest of the body. Rarely limited pattern of pale brown spots on the tail. The black metatarsal tubercles already appear before full development of the posterior limbs.
Bioacoustics. The amplexus call of a single P. varaldii male (27 notes) and female (34 notes) originating from Forêt de Mamora were analysed and presented here for the first time ( Fig. 7E&F View FIGURE 7 ). Calls were emitted at regular intervals in trains of up to seven notes. The notes were usually composed of three different pulses, of which the first two pulses are weak and brief whereas the third pulse is more powerful, longer and consists of a train of very short pulses. Occasionally, the first note is only composed of a single pulse (6.1% of analysed notes). Note length ranged between 52 ms to 94 ms (averagely 85 ± 09 ms) in case of the male, and between 60 ms and 91 ms (averagely 83 ± 06 ms) for the female. The dominant frequency ranged from 589 to 1370 Hz for the male and from 675 to 981 Hz for the female. Compared to P. cultripes , the amplexus call of P. varaldii has a higher per-note pulse number and a higher frequency ( Lizana et al. 1994).
As reviewed by Lizana et al. (1994), Pelobates sp. are characterized by a rich vocal repertoire, whereas the advertisement call and release call of P. varaldii remain undescribed. Further study is therefore required in order to describe and assess the degree of differentiation between the calls of P. varaldii and other members of the genus.
Natural history. Pelobates varaldii is the most stenoecious Moroccan amphibian species, of which the occurrence is limited to the vicinity of Mediterranean temporary ponds located on loose, sandy soils at low altitudes. Additionally, the presence of significant tree cover ( Quercus suber forests, although planted Pinus , Acacia and Eucalyptes forests are also occupied) seems to be of high importance ( de Pous et al. 2012). Pelobates varaldii can be extremely common within suitable habitat; de Pous et al. (2012) encountered the species in 95% of more than 150 ponds located in and around Forêt de Mamora near Rabat. Natural history of the southernmost populations on the stony Doukkala plain (i.e. near Oualidia, Crochet & Geniez 2003) considerably differs, with individuals being highly localized, occupying fragmented, largely barren areas of loose sandy soil ( de Pous et al. 2012).
Guarino et al. (2011) analysed age structure and growth of a population located in Forêt de Mamora. The age of the analysed individuals ranged between 2–7 years in males (mean 4.5±1.2, n = 66) and 2–10 years in females (4.7 ± 2.4, n = 20). Pelobates varaldii aestivates in the soil from late spring until the first winter rains. Activity is strictly nocturnal during the reproductive period, although aquatic individuals might remain day-active for several days during the activity peak (P. de Pous & W. Beukema pers. obs.). Reproduction takes place from the end of October to the end of January, with larvae being observed between November and April ( El Hamoumi 1988; El Hamoumi & Himmi 2010; Lapeña et al. 2011; P. de Pous pers. obs.). Recently metamorphosed juveniles have been observed in May and June ( Pasteur & Bons 1959). General activity continues up to May ( Crochet & Geniez 2003; Guarino et al. 2011).
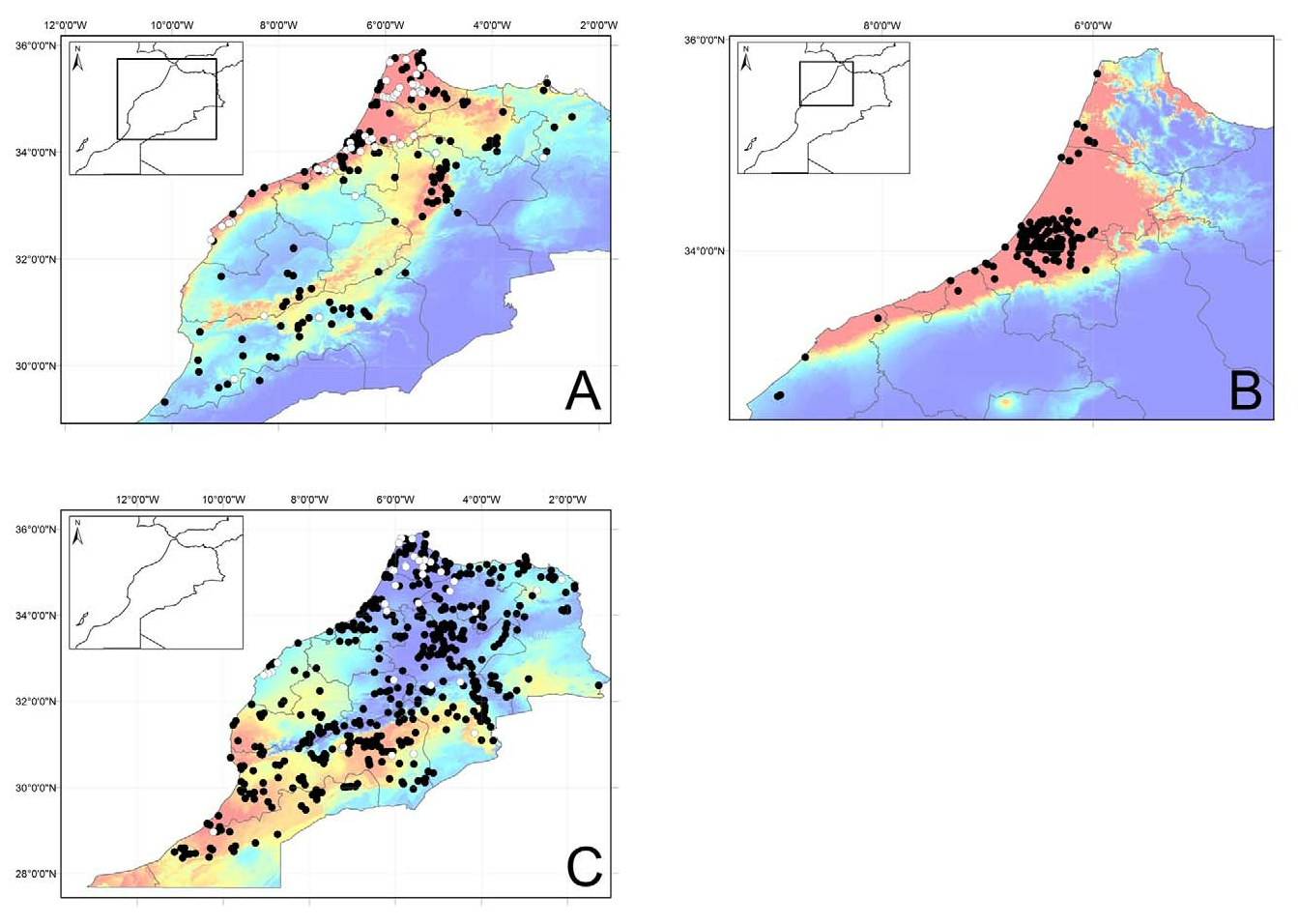
Distribution. Distribution data on this cryptic species were limited until recently, notwithstanding the contributions of Crochet and Geniez (2003), Guzman et al. (2007) and El Hamoumi and Himmi (2010). The description of abundant calling P. varaldii in the High Atlas Mountains by Malkmus (1983) should be considered erroneous. Lapeña et al. (2011) provided a record from the northern Moroccan coast between Asilah and Tanger, thereby extending the distribution some 50 kilometres to the north, confirming predictions made by Pasteur and Bons (1959). de Pous et al. (2012) provided a detailed distributional review and threat assessment of the species, confirming the occurrence of P. varaldii in four isolated areas along the Atlantic coast, of which at least two are highly threatened. The distribution map ( Fig. 16B View FIGURE 16 ) is composed of records from Bons and Geniez (1996), Crochet and Geniez (2003), Guzman et al. (2007), El Hamoumi and Himmi (2010), Lapeña et al. (2011) and de Pous et al. (2012).
National Red List Status. Endangered.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.