
Desoria calderonis Valle, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.787.1599 |
|
publication LSID |
lsid:zoobank.org:pub:0C6C4FA5-D1D8-4C2F-825F-A3882DE02B99 |
|
DOI |
https://doi.org/10.5281/zenodo.5837952 |
|
persistent identifier |
https://treatment.plazi.org/id/1DEC543F-66C5-4556-B5B3-6806CEB35488 |
|
taxon LSID |
lsid:zoobank.org:act:1DEC543F-66C5-4556-B5B3-6806CEB35488 |
|
treatment provided by |
Felipe |
|
scientific name |
Desoria calderonis Valle |
| status |
sp. nov. |
Desoria calderonis Valle View in CoL sp. nov.
urn:lsid:zoobank.org:act:1DEC543F-66C5-4556-B5B3-6806CEB35488
Figs 1–5 View Fig View Fig View Fig View Fig View Fig
Diagnosis
Desoria calderonis sp. nov. belongs to the violacea -group sensu Potapov, 2001, with quadridentate mucro without seta, maxillary palp bifurcate, apical folds on labrum sharp. Differences from closely related species are analysed in the section ‘Taxonomic and ecological consideration’.
Etymology
The epithet of the new species reflects the name of the site from which holotype and paratypes derive, Calderone glacier ( Italy, Abruzzo, Gran Sasso massif).
Material examined
Holotype ITALY • ♀; Abruzzo (Central Italy), Gran Sasso massif, Apennines , Calderone Glacier , supraglacial stony debris; 42°28ʹ16.2″ N, 13°34ʹ05.8″ E; alt. 2700 m a.s.l.; 8 Jul. 2020; B. Valle and M. Di Musciano leg.; collected with flotation method; Genbank ( NCBI) MZ686962 View Materials -70 ; Collembola collection, Department of Life Sciences , University of Siena , Italy. GoogleMaps
Paratypes ITALY • 15 spec. (♂ and ♀); same collection data as for holotype; Collembola collection, Department of Life Sciences , University of Siena, Italy GoogleMaps .
Description
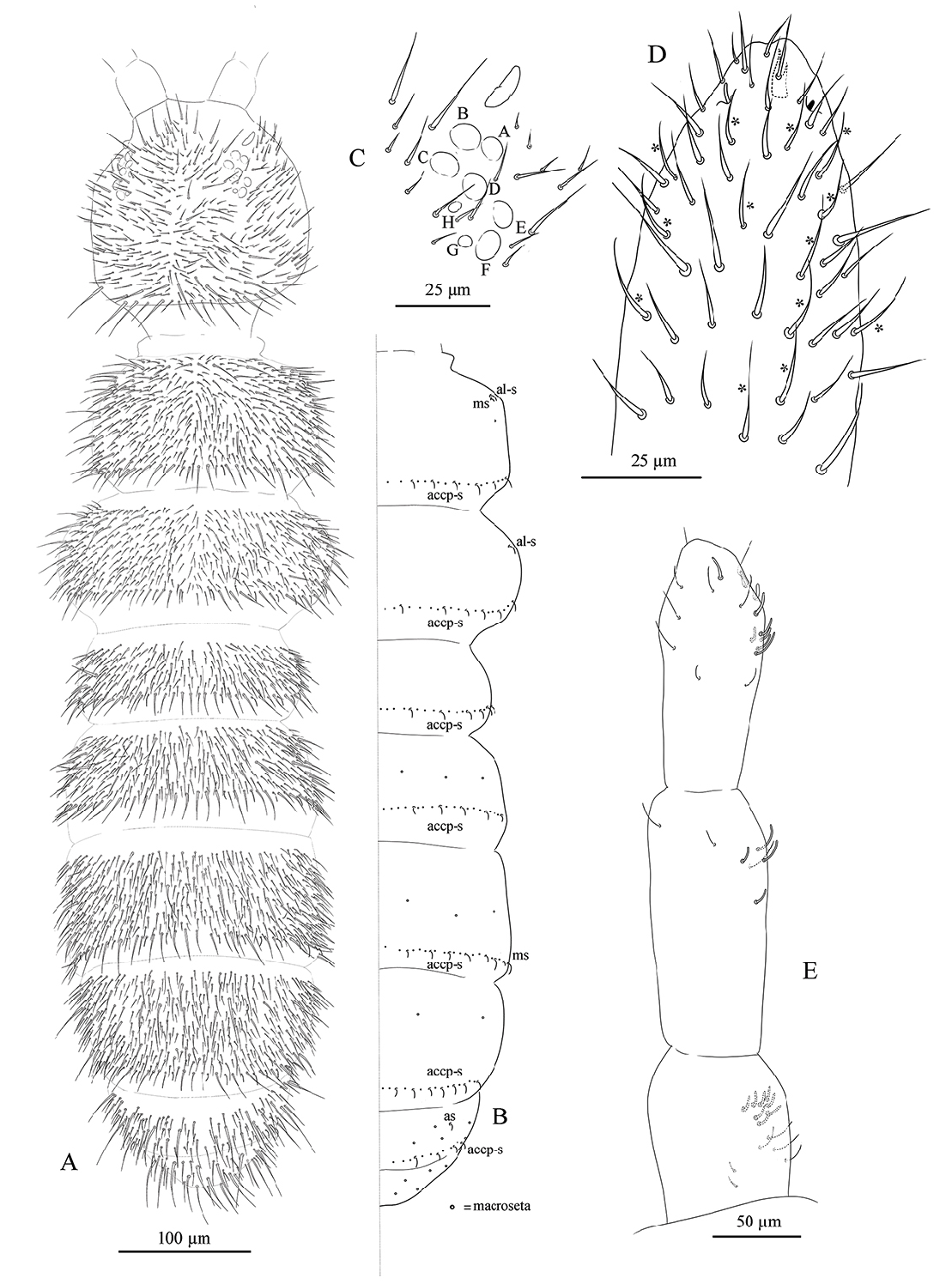
BODY. Mean body length. 1.5 mm (standard deviation: 0.1 mm on 12 specimens, see Table 1 View Table 1 ). Colour violet-black on abdomen and antennae, lighter on furca and legs, which are brownish ( Fig. 1 View Fig ); juveniles are much paler, bluish. Cuticle granulation fine and regularly distributed; all dorsal tergites clearly separated from each other. Abd. III and IV of approximately same width.
CHAETOTAXY. Terga plurichaetotic, consisting of micro-, meso- and macrosetae, these latter well differentiated on last abdominal tergites (Abd. IV–VI, in median position), but not well distinguished from ordinary setae on other tergites ( Fig. 2A View Fig ). On every segment, longer setae concentrated along the lateral and posterior edges of tergite. All setae smooth. Macrosetae on Abd. V 0.9–1 times median length of tergite and 1.8–2.4 times as long as inner edge of Claw III ( Table 1 View Table 1 ). Sensory chaetotaxy constituted by ms -setae, accp-, al- and as- setae. Only Th. II and Abd. III have ms- setae (formula 10/001). Dorsal s -setae constituted by single al-s eta on Th. II and Th. III, single as -seta on Abd. V and by accp -setae (4–7 on each tergite from Th. II to Abd. V) normally set within p -row ( Fig. 2B View Fig ). The number of accp -setae can be expressed as 5–6,6/5–6,5,7,6–7,4 ( Fig. 2B View Fig ). The number of ordinary setae between accp- setae may vary by 1 (rarely 2) with respect to the scheme presented in Figure 2B View Fig (after Potapov 1989).
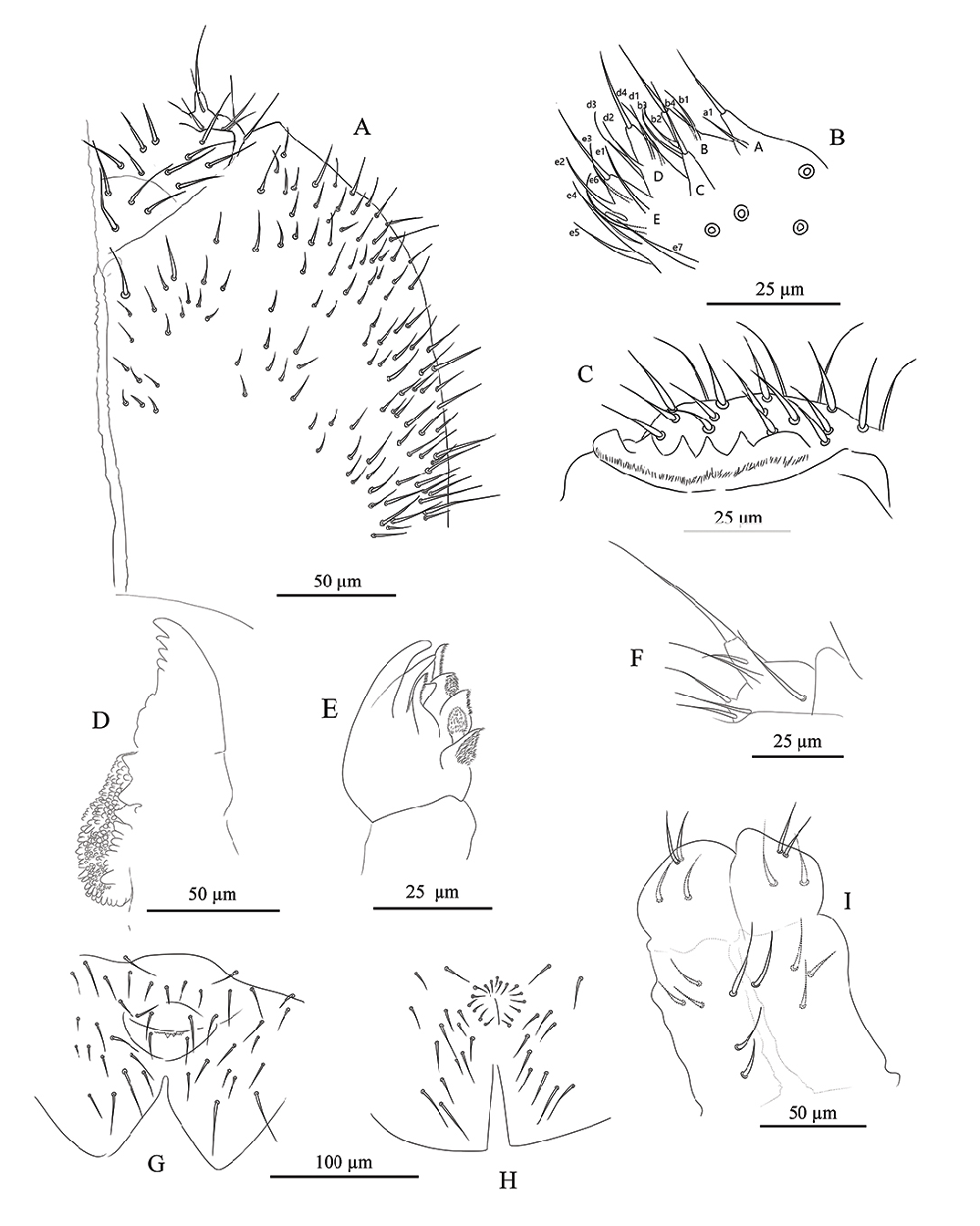
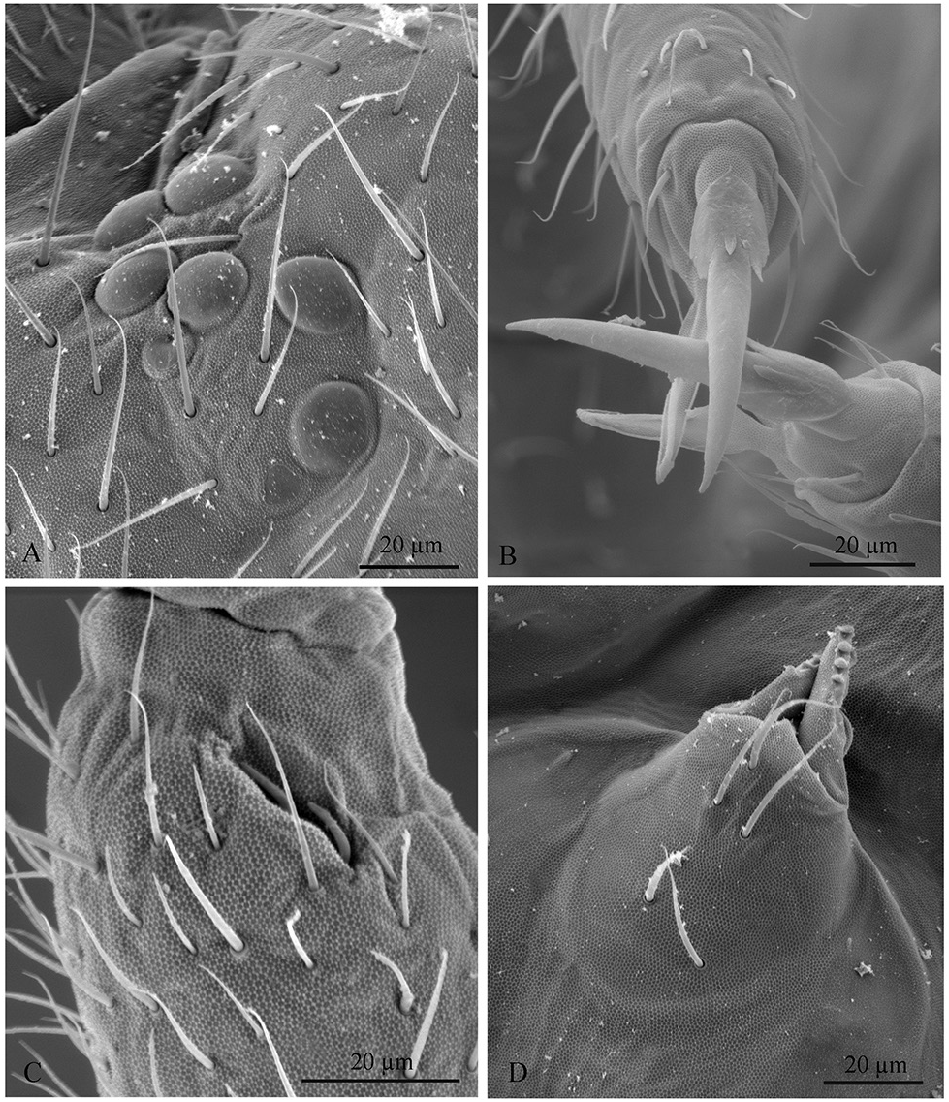
HEAD. Antennae longer than cephalic diagonal (D/A = 0.79). Ratio among Ant. I/Ant. II/Ant. III/Ant. IV is 1/1.75/1.57/2.57. Some s -setae well differentiated, others thick and hardly differing from ordinary setae (hereafter ʻseta-like s -setaeʼ). There are often cases of asymmetry among s -setae between antennae of the same specimen. Ant. I has about 54 setae, 6–11 short, thick and cylindrical s -setae and 4 seta-like s -setae in ventro-lateral position; 2–3 microsetae in ventro-proximal position ( Fig. 2E View Fig ). Ant. II has about 90 setae, 4 s -setae and 2–3 seta-like s -setae ( Fig. 2E View Fig ). Ant. III has about 84 setae and a sensory field that includes 2 s -setae of AO III, about 8 s -setae and 6 seta-like s -setae ( Figs 2E View Fig ; 5C View Fig ). Ant. IV plurichaetotic with more than 300 setae, with few s -setae and several seta-like s -setae; one simple small subapical, rod-shaped organite and a clearly bifurcate pin-like seta ( Fig. 2D View Fig ). Eye spots strongly dark pigmented with 8 + 8 ocelli (G and H usually hardly visible; Figs 2C View Fig , 5A View Fig ). PAO elongated, with a weak median constriction, about 2 times as long as diameter of nearest ocellus ( Fig. 2C View Fig ). Prelabral setae 4. Labral formula as 5, 5, 4 and 4 sharp papillae ( Fig. 4C View Fig ). Maxillary palp bifurcate and maxillary outer lobe with 4 sublobal hairs ( Fig. 4F View Fig ). Labial palp with 5 papillae and a total of 16 guard setae ( Fjellberg 1999) distributed as: A 1, B 1–4, C 0, D 1–4, E 1–7 ( Fig. 4B View Fig ). Hypostomal papilla with H as long as h 1 /h 2. Proximal (px), basomedian (bm) and basolateral (bl) fields of labium with 4, 4 and 5 setae, respectively ( Fig. 4A View Fig ). Maxilla of normal shape as in Fig. 4E View Fig , with lamellae shorter than capitulum. Ventral line of head with 9 + 9 postlabial setae ( Fig. 4A View Fig ). Mandible with well-developed molar plate as in Fig. 4D View Fig . VT with 2–4 + 2–4 anterior, 4 + 4 latero-distal and 4 posterior setae with 2 in apical transverse row ( Fig. 4I View Fig ). Retinaculum with 4 teeth and 7–8 setae ( Figs 3F View Fig , 5D View Fig ).
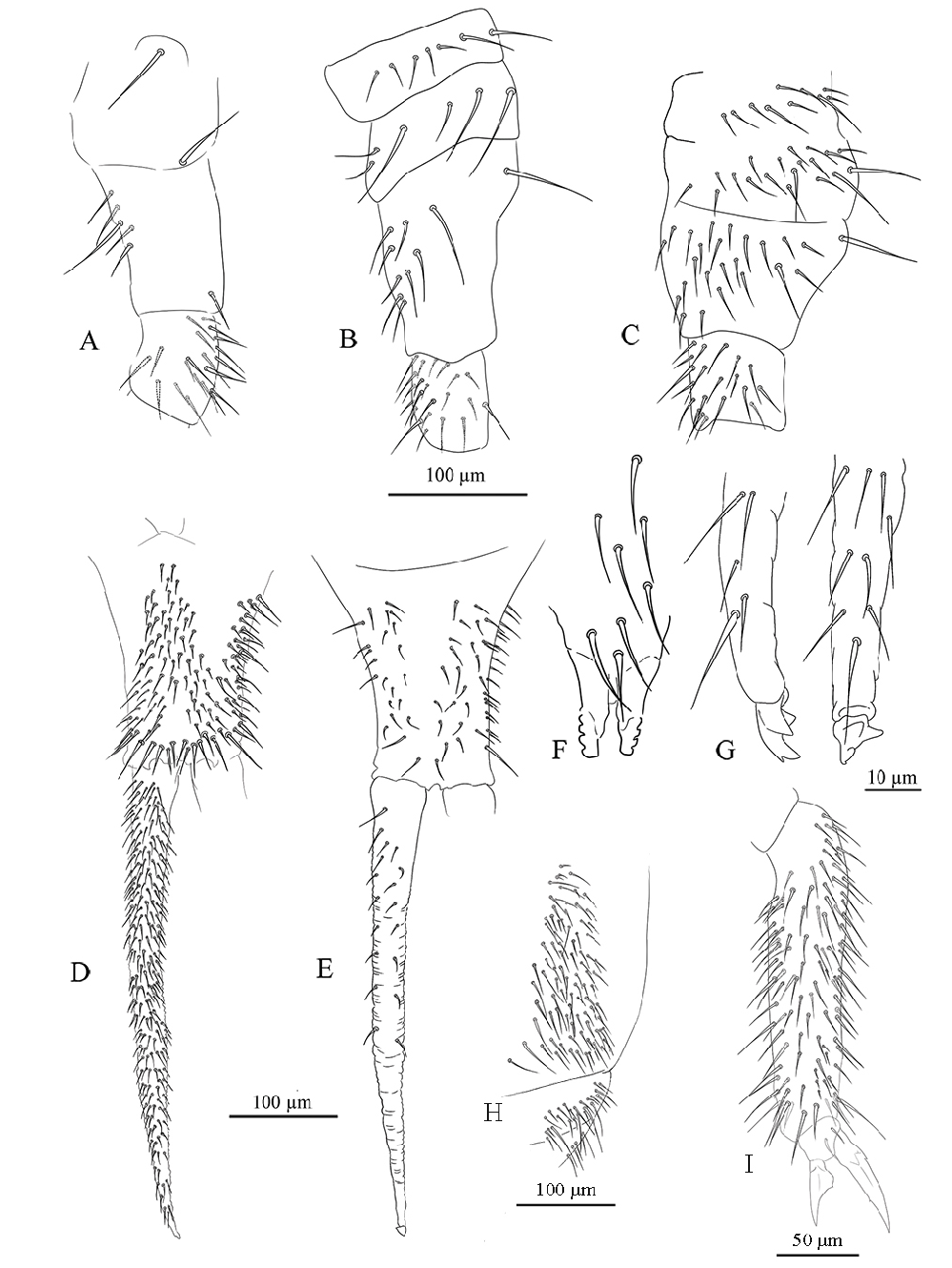
FURCA. Well-developed; ratio of mucro/dens/manubrium = 1/41/19 ( Fig. 3D–E View Fig ). Ventral setae on manubrium numerous (about 88) and ventro-apical setae (10–12) larger than the others ( Fig. 3D View Fig ), with the exception of 2 + 2 short apical setae; more than 70 dorsal setae ( Fig. 3E View Fig ). Dens with dorsal crenulations, about 200 ventral and 18 dorsal setae ( Fig. 3D–E View Fig ). Mucro quadridentate with apical tooth much smaller than subapical one ( Fig. 3G View Fig ).
LEGS. Upper and lower subcoxa of Leg I with 1 outer seta ( Fig. 3A View Fig ). Upper subcoxa of Leg II with 7, lower subcoxa with 6 outer setae ( Fig. 3B View Fig ). Upper subcoxa of Leg III with 10, lower subcoxa with 19 outer setae ( Fig. 3C View Fig ). Coxa with 5, 10, 24 setae, respectively, on leg I, II, III ( Fig. 3A–C View Fig ). Trochanter with 20, 21, 20 setae, respectively, on leg I, II, III. Femur with 36, 42, 74, respectively, on leg I, II, III. Tita with 45, 54,96 setae, respectively, on Leg I, II, III; tenent hair pointed ( Fig. 3I View Fig ). Claw of normal shape with lateral and inner teeth; empodium with a small inner tooth; pretarsus with a pair of setae ( Fig. 3I View Fig ).
Measurements
See Table 1 View Table 1 .
Ecology
Desoria calderonis sp. nov. was found only on supraglacial stony debris of the Calderone glacier at 2650–2700 m a.s.l. (Appendix 1A). The supraglacial stony debris (carbonate: dolomite and limestone) is very coarse and the fine component is poorly represented (Appendix 1B). In this mineral, inorganic soil, D. calderonis sp. nov. was found mostly where the debris is in contact with the ice (Appendix 1B). In this environment, the vegetation cover is almost absent, with the rare exception of sporadic seedlings of Arabis alpina ssp. caucasica (Willd.) Briq. On supraglacial debris the mean annual temperature is 0.67°C (4.6°C during the snow-free period), the minimum recorded was -7.7°C and maximum 36.1°C; the snow cover persists on average for 250 days a year; the relative humidity is on average 95.7% during the whole year (85.9% during the snow-free period). Since D. calderonis sp. nov. was found only on supraglacial stony debris, and not in the surrounding habitats (LIA moraines: mean annual temperature = 1.3°C, mean temperature during snow-free period = 4.4°C; minimum = -9.8°C; maximum = 40.7°C; annual relative humidity = 88.7%; mean relative humidity during the snow-free period = 84.7°C; snow cover duration = 146 days), we can consider this species as cryophilic. On supraglacial debris, pH is 8.6, the organic matter content is 1.7 g /kg and carbonate calcium content is 96.2%, while on LIA moraine, pH is 8.3, organic matter content is 3.9 g /kg and carbonate calcium content is 91.4%.
Distribution
Desoria calderonis sp. nov. is currently known only for the type locality.
Type locality
Gran Sasso massif, Apennines, Calderone glacier (42°28ʹ16.2″ N, 13°34ʹ05.8″ E). Supraglacial stony debris of Calderone glacier, altitude: 2650–2700 m a.s.l.
Taxonomic and ecological considerations
Desoria calderonis sp. nov. belongs to the violacea -group sensu Potapov 2001, having mucro quadridentate, without seta, maxillary palp bifurcate, apical folds on labrum sharp. It differs from most species of this group ( Table 2 View Table 2 ) by the reduced number of setae on VT (with the exception of D. duodecemoculata (Denis, 1927) and D. nivalis (Carl, 1910) , which also have few setae, but in a different number). Desoria calderonis sp. nov., in addition, has a characteristic sensory field on Ant I with short, thick and cylindrical s -setae; this characteristic is common, for Palearctic Desoria , to many species of the fennica -group (sensu Potapov 2001) – D.atkasukiensis (Fjellberg, 1978) , D. fennica (Reuter, 1895) sensu Fjellberg 1979 , D. fjellbergi ( Najt, 1981) , D. iuxta (Dunger, 1982) , D. kaszabi (Dunger, 1982) , D. saltans , D. tigrina Nicolet, 1842 – two species of olivacea -group – D. infuscata (Murphy, 1959) and D. olivacea (Tullberg, 1871) sensu Fjellberg 1979 – and only one species of the violacea- group – D. nivea (Sch ӓffer, 1896). Among species of the violacea -group, another peculiarity of D. calderonis sp. nov. is the number of dorsal setae on the dens, similar only to that in D. neglecta (Sch ӓffer, 1900) sensu Fjellberg 1978, D. taimyrica (Martynova, 1974) and D. violacea (Tullberg, 1876) sensu Fjellberg 1979 . In general, D. calderonis sp. nov. differs by a combination of at least four important characters ( Table 2 View Table 2 ) from every species of the group.
Within the group, considering the number of common characteristics, D. calderonis sp. nov. appears most similar to D. duodecemoculata – present in Italy, Austria, Spain and France ( Potapov 2001) – and D. nivalis , present in the Alps ( France, Switzerland, Austria) and possibly in eastern Europe ( Potapov 2001) ( Table 2 View Table 2 ). Nevertheless, some features allow us to discriminate the new species from these. First, D. duodecemoculata and D. nivalis do not have short, thick and cylindrical, but only hairlike s -setae on Ant I. In addition, the new species differs from D. duodecemoculata by the chaetotaxy of VT and the number of dorsal setae on the dens; it differs from D. nivalis by having Ant II-III violet-black (white in D. nivalis ), a longer PAO and by the number of dorsal setae on the dens.
Both D. duodecemoculata and D. nivalis belong to the nivalis -complex, a group of European mountain species included in the violacea -group. Desoria nivalis , in particular, is known to live near snow fields and other cold sites in high mountains ( Handschin 1924; Franz & Serrl-Butschek 1954), while the ecology and the taxonomy of the other members of the complex need to be revised ( Potapov 2001). Because of these similarities in morphology and ecology, we could ascribe our species to this nivaliscomplex, even if we reported marked differences, in particular the presence in D. calderonis sp. nov. of the sensory field on Ant. I.
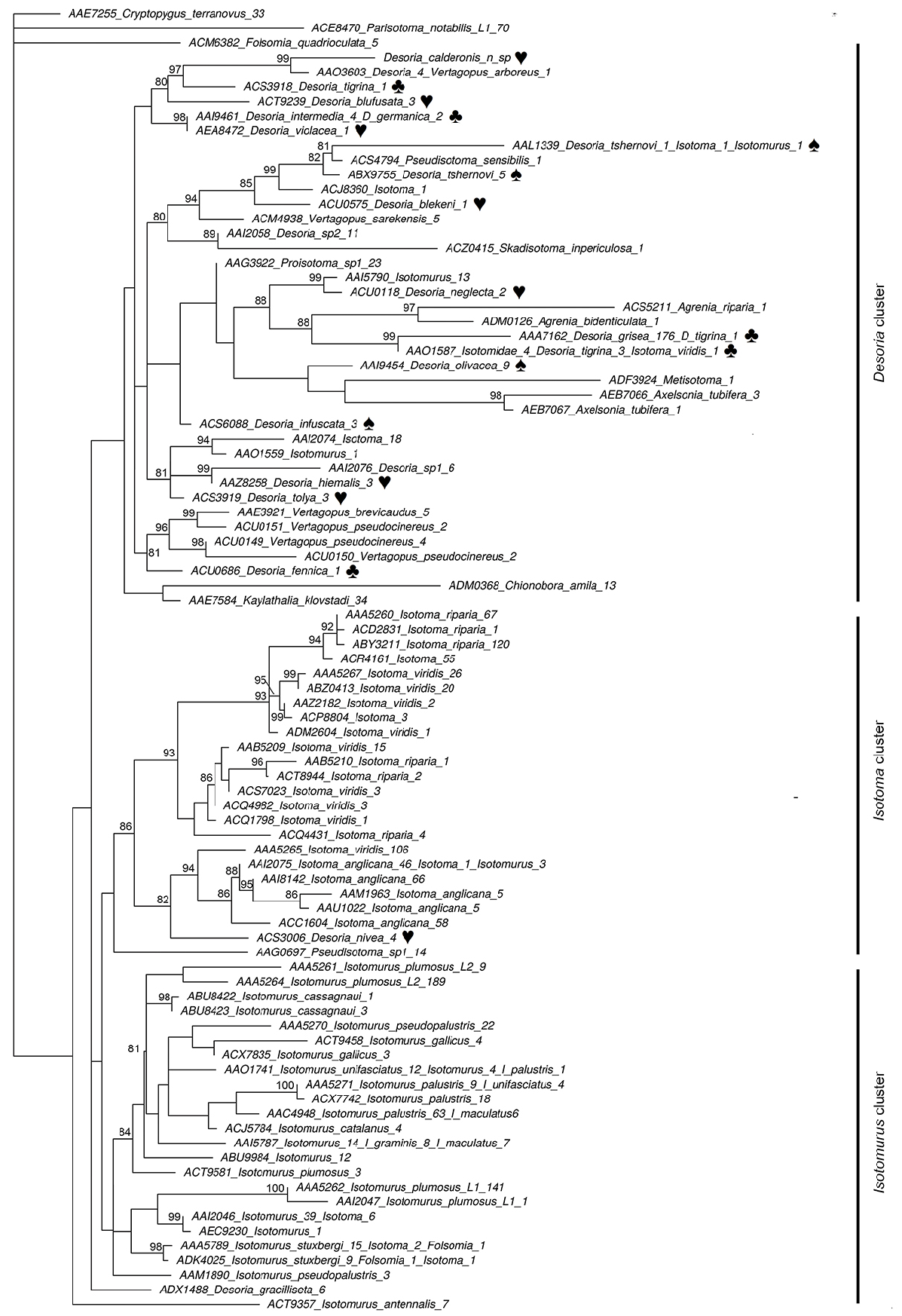
Phylogenetic context
The phylogenetic tree ( Fig. 6 View Fig ), with a log-likelihood of 3077.45, is characterized by good support at recent nodes but low support at deeper nodes. It appears subdivided into three major clusters, two dominated by Isotoma and Isotomurus , respectively, and one by Desoria , with records representing 14 different Desoria species as well as others incompletely identified as Desoria sp. This latter cluster also included scattered sequences from the following genera: Vertagopus , Pseudisotoma , Isotoma , Skadisotoma , Proisotoma , Isotomurus , Agrenia , Metisotoma , Axelsonia , Chionobora and Kaylathalia . Within the Desoria cluster, Desoria calderonis sp. nov. clustered with high support (99) with one bin (AAO3603) identified as Desoria (with no indication on the species, 4 sequences from France) and Vertagopus arboreus (1 sequence from Ontario, Canada). The mean divergence between D. calderonis sp. nov. and this bin is 16.9% (S.D. 0.2). Both bins were associated (support 97) with one record of D. tigrina and this latter cluster, although with lower support, was associated with records of D. blufusata , D. germanica , D. intermedia and D. violacea .
Apart from D. violacea (BOLD:AEA8472), D. tigrina (BOLD:ACS3918), Vertagopus arboreus (BOLD:AAO3603), and individual records mined from GenBank with no location information, all others reports come from areas characterized by a markedly cold climate, mostly in Northern Europe. Similarly to D. calderonis sp. nov., D. blufusata (BOLD:ACT9239, 3 records), D. intermedia (BOLD:AAI9461, 4 records) and D. germanica (BOLD:AAI9461, 2 records; see Stevens et al. 2006 for sampling information) were collected in low-temperature habitats, specifically in Norway and Sweden, with two records of D. intermedia explicitly associated to a snow covered setting. At variance, Desoria sp. (BOLD:AAO3603), for which species level identification is missing, originated from British Columbia ( Canada) as well and the Ile-de-France ( France).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Isotominae |
|
Genus |