
Cacosternum nanogularum, Channing, Alan, Schmitz, Andreas, Burger, Marius & Kielgast, Jos, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3701.5.2 |
|
publication LSID |
lsid:zoobank.org:pub:44F12E00-E270-41B0-ADE9-4FF774C975C3 |
|
DOI |
https://doi.org/10.5281/zenodo.5663615 |
|
persistent identifier |
https://treatment.plazi.org/id/A93A410D-6E4B-FFAF-FF03-A38EDB0DD4CC |
|
treatment provided by |
Plazi |
|
scientific name |
Cacosternum nanogularum |
| status |
sp. nov. |
Cacosternum nanogularum View in CoL sp. nov.
Small-throated Dainty Frog. ( Figs. 3 View FIGURE 3 I, 4I)
Holotype. An adult male MHNG 2740.77 collected by M. Burger, 3 December 2005, on the road between Eshowe and Nkandla, KwaZulu-Natal Province, South Africa, 28° 51’ 04” S, 31° 12’ 27” E. The specimen was collected from a roadside puddle.
Paratypes. Six males; MHNG 2740.75 from 41 km from Kranskop on road to Qudeni, KwaZulu-Natal Province, South Africa, 28° 43’ 41” S, 30° 56’ 35” E; NMB A7518–20, MHNG 2740.76 from 13 km from Kranskop along road to Qudeni, KwaZulu-Natal Province, South Africa, 28° 54’ 14” S, 30° 51’ 15” E; MHNG 2740.78 collected on the road between Eshowe and Nkandla, KwaZulu-Natal Province, South Africa, 28° 51’ 04” S, 31° 12’ 27” E; a female, MHNG 2740.74, from Dukuduku, KwaZulu-Natal Province, South Africa, 28° 21' 09" S, 32° 17' 53" E.
Diagnosis. The uncorrected p distances for 16S from the other 14 species ranges from 3.8–6.9%, while the within-clade p distances range from 0–0.4 (n=7). The sample has five likely tyr haplotypes of which none are shared. This species lacks a distinct supratympanic fold, which is present in all the other species. It can be distinguished from C. capense by the absence of large lateral and posterior glands on the dorsum; from C. karooicum which has the nostrils at least three nostril diameters back from the snout margin; from C. leleupi which has a small amount of webbing; from C. namaquense which has a diffuse grey vocal sac anteriorly (dark in C. nanogularum sp. nov.) ( Fig. 4 View FIGURE 4 ). It has three supernumerary tubercles on the palm, less than the five of C. aggestum sp. nov. and C. leleupi . The inner metatarsal tubercle is equal to the proximal subarticular tubercle of the first toe, differing from the wider inner metatarsal tubercle of C. aggestum sp. nov., C. boettgeri and C. parvum . The rictal gland is indistinct or absent, which distinguishes it from the prominent glands of C. aggestum sp. nov., C. boettgeri , C. australis sp. nov., C. striatum , C. nanum , C. parvum , C. platys , C. kinangopensis and C. plimptoni . The advertisement call is a series of chirps, which distinguishes it from the regular single or multiple clicks of C. boettgeri , C. kinangopensis , C. leleupi , C. plimptoni and the brief chirp of C. nanum . It is unlike the 'bouncing marble' calls that consist of a number of pulses that speed up, as C. aggestum sp. nov., C. australis sp. nov. and C. platys , or the long complex call of C. rhythmum sp. nov. The call is not a creak as in C. capense , C. karooicum , C. namaquense , C. parvum , or C. striatum .
Description of the holotype. The holotype is an adult male, SUL 19.7. Body widest at mid-belly, with a narrow head (HW/SUL 0.33). The head is bluntly rounded from above and in profile. Head length measured from the angle of the jaw is about one third of body length (HL/SUL 0.31). Canthus rostralis rounded, straight from eye to nostril, loreal region sloped outwards ventrally; nostrils small, rounded, directed laterally, and posteriorly. The nostrils are placed nearly midway between the snout than the eye (EN/SL 0.52). Internostril distance is greater than distance between eye and nostril (NN/EN 1.31). Eyes directed anterolaterally, the eyes protrude, and are visible from below, relatively small (ED/HW 0.35; ED/SUL 0.12), less than snout (ED/SL 0.74). Distance between anterior corners of eyes subequal to internostril distance (NN/EE 0.53). The angle of the jaw is posterior to a line drawn vertically from the back of the eye. The tympanum is not visible. Jaws without dentition; choanae small, round, located at anterior margins of roof of mouth; vomer processes and teeth absent; tongue long, narrow, slightly bifurcated distally. No median lingual papilla present.
The dorsal surfaces of the head, trunk and limbs are smooth, with glands and skin folds present but indistinct; the rictal gland is smooth, continuing posteriorly as a series of bulges to the arm insertion. The supratympanic fold forms a forms a groove running over the arm to the leg insertion. Lines of broken skin glands form an hourglass pattern dorsally, narrowest at the scapular region. The underside is smooth, with a slightly pigmented vocal sac with lateral folds.
The fore limb is slender, hand small (HAN/SUL 0.30), finger tips bluntly rounded without discs. Relative finger lengths I<II<IV<III; subarticular tubercles distinct, rounded, with one on fingers I and II, two on fingers III and IV. No webbing between fingers. Thenar tubercle small, rounded, partially obscured by nuptial pad that reaches the distal phalanx of the first finger; palmar tubercles and inner metatarsal tubercle moderate, rounded, outer metacarpal tubercle present. There is a single supernumerary tubercle on the palm.
Hind limbs moderately long (TIB/SUL 0.50; FOT/SUL 0.52), foot subequal to tibia (TIB/FOT 0.95); thighs are moderately developed, with rough glands on the inner posterior faces; relative toe lengths are I<II<V<III<IV. The toe tips are not expanded; subarticular tubercles: one on toes I and II, two on toes III and V, and three on toe IV. Only a trace of webbing between the toes. Inner metatarsal tubercle conical, prominent, outer metatarsal tubercle present as a small pale raised spot. Morphological proportions are shown in Appendix 2.
Colour in life. The background colour is beige, overlain with a dark brown interorbital bar with a sharp tip pointing posteriorly, and a black band running from the nostril, through the eye to the arm insertion. The upper lip is white with black speckles, continuing as a white rictal gland. Colour in preservative. The patterns are visible as dark browns on a paler background.
Paratype variation. The male paratypes have SUL 20.4–22.5, with the females 23.2–25.0. They are very similar to the holotype in having discontinuous bumpy rictal glands, large subarticular tubercles on the hand, and small conical subarticular tubercles at the base of the fourth toe. The belly spots are similar, with the female throats spotted but less pigmented than the males, while the body proportions show little variation.
Advertisement call. The general call structure consists of a series (10 to>20) of regularly repeated pulsed notes (chirps), with a note rate of 0.4– 0.6 s -1. The mean note duration is 89 ms, with the mean number of pulses per call of 9. The mean pulse rate is 89 s -1. It has a mean emphasised frequency of 3.02 kHz. The illustrated call ( Fig. 10 View FIGURE 10 ) has a duration of 0.1 s, with 9 pulses at a pulse rate of 80 s -1.
Eggs and tadpoles. Unknown.
Distribution. Molecular samples come from Nkandla and the Lebombo Mountains, with additional specimens known from Qudeni and Dukuduku ( Fig. 9 View FIGURE 9 ). The full extent of its range is unknown. The frogs were found in flooded temporary pools with emergent vegetation.
Etymology. The specific name is derived from the Latin nanus (small), and gula (throat). It is an adjective. The name refers to the very small vocal sac that is produced during calling.
Cacosternum poyntoni Lambiris, 1988
Genetics. The uncorrected p distances between this species and the other 14 range from 2.8–6.0%, while the within-clade p distances range from 0.0–0.4 (n=20). The sample has 16 likely tyr haplotypes, of which one is shared between a single C. nanum and one C. rhythmum sp. nov.
Advertisement call. Based on an analysis of 447 calls of 41 specimens, a calling bout usually consists of a long (15 to>30) series of pulsed chirps that are rapidly repeated, about 1.2 s -1 (range 0.6– 2.0 s -1). The mean call (chirp) duration is 52.6 ms (range 19.5–99.7 ms). The mean number of pulses per call is 11 (range 5–18), with a mean pulse rate of 200 s -1 (range 95–288 s -1). The mean emphasised frequency is 3.6 kHz (range 3.3–4.1 kHz) A spectrogram of a typical call is shown in Fig. 10 View FIGURE 10 . It has a duration of 0.04 s, consisting of 11 pulses at a pulse rate of 250 s -1.
Morphology. Females reach 18 mm SUL. The dorsum lacks protruding glands. Viewed from above, the nostrils are close to the anterior margin of the snout. The supratympanic fold continues as a straight line from eyelid to arm insertion. The rictal gland is interrupted, tapering to the arm insertion. There are four supernumerary tubercles on the palm. The inner metatarsal tubercle is low, with a width equal to the tip of the first toe. Breeding males possess an inconspicuous nuptial pad that extends to the last phalanx. The belly spots are the size of the lens of the eye, dense grey, often running together ( Fig. 3 View FIGURE 3 ).
Distribution. This species is known from the south and east of South Africa, occurring in 276 quarter degree squares (Animal Demography Unit 2012). Molecular samples come from Baviaanskloof, Cedarville, Goukamma, Hogsback, Kubusi Forest Reserve, Maclear and Montague Pass in South Africa ( Fig. 7 View FIGURE 7 ). It breeds in a variety of water bodies (Scott 2004b), usually those with emergent vegetation.
Note. We regard Cacosternum poyntoni as a junior synonym of C. nanum , following the reasons presented by Minter (2004), Scott (2004b) and Scott & Minter (2004a, b). These include the lack of material apart from the type, after nearly 60 years, from a well-known locality, Pietermaritzburg; the consensus amongst local herpetologists that this is not a valid taxon; and the skeletal similarity to C. nanum from an unpublished report.
Cacosternum parvum Poynton, 1963 Mountain Dainty Frog.
( Figs. 3 View FIGURE 3 K, 4K)
Genetics. The within-clade variation in uncorrected p distances for the 16S fragment is 0.2%, with the differences to the other 14 species being 3.9–7.6%. The sample has two tyr haplotypes, of which none are shared.
Advertisement call. The call is a brief chirp. The following description is based on an analysis of 172 calls of 17 specimens. A motivated calling bout usually consists of a long (10 to>20) series of pulsed chirps that are repeated at a rate of about 0.6 s -1 (range 0.4– 0.7 s -1). The mean call (chirp) duration is 60.2 ms, (range 27.6–109.0 ms). The mean number of pulses per call is 6 (range 3–9) with a mean pulse rate of 86.8 s -1 (range 59.5–125 s -1). The mean emphasised frequency is 3.99 (range 3.55–4.42 kHz). The illustrated call ( Fig. 10 View FIGURE 10 ) has a duration of 0.06 s, with seven almost equally spaced pulses, with a pulse rate of 100 s -1.
Morphology. Females do not exceed 16.5 mm SUL in our sample. Breeding males have dark vocal sacs with a paler margin. The dorsum is smooth, and, viewed from above the nostrils are positioned on the anterior edge of the snout. The supratympanic fold continues posteriorly to form a rounded saddle. The rictal gland is indistinct, interrupted. There are no supernumerary tubercles on the palm. There is no webbing, with the notch between the third and fourth toes reaching the proximal subarticular tubercle of the third toe. The outer metatarsal is absent or indistinct, while the inner metatarsal tubercle is low, twice the width of the tip of the first toe. The nuptial pad in breeding males is slightly pigmented but indistinct. The belly markings consist of groupings of brown speckles that form blotches.
Distribution. Molecular samples come from Mariepskop ( Fig. 9 View FIGURE 9 ). The species appears to be a highland endemic in eastern South Africa (Scott & Minter 2004b), where it breeds in high altitude grassland pools (Scott 2004e).
Note. The GenBank sequence DQ 022353 View Materials with the voucher identified as Cacosternum nanum parvum (Scott 2005) is a C. nanum sequence.
Cacosternum platys Rose, 1950 Flat Dainty Frog.
( Figs. 3 View FIGURE 3 L, 4L)
Genetics. The within-clade variation in uncorrected p distance for the 16S fragment varies from 0.0–0.2%, but between C. platys and the other 14 species 1.8–6.6%. The sample has nine likely tyr haplotypes, of which one is shared with a specimen of C. australis sp. nov. See the Discussion.
Advertisement call. A typical advertisement call ( Fig. 11 View FIGURE 11 ) consists of a long series of pulses that increase in tempo. The duration of the illustrated call is 0.55 s, with 34 pulses. The initial and final pulse rates are 20 s -1 and 80-1.
Morphology. This is a small species, with females less than 14 mm in our sample. The vocal sac in breeding males is pale grey ( Fig. 4 View FIGURE 4 ). The dorsum is smooth. Viewed from above, the nostrils are situated on the anterior margin of the snout. The supratympanic fold continues posteriorly in a straight line from the eyelid to the arm insertion. The rictal gland is interrupted. There are two supernumerary tubercles on the palm. There is no webbing. The inner metatarsal tubercle is low, rounded, equal in width to the tip of the first toe. The nuptial pad is a dark swollen mass at the base of the first finger, tapering to the level of the proximal tubercle. The belly pattern consists of very small spots consisting of groups of speckles.
Distribution. Molecular evidence shows the species is present on the Cape Peninsula (Noordhoek) and in Cape Town (Kenilworth) ( Fig. 7 View FIGURE 7 ). It is a fynbos species, breeding in low altitude temporary pools. Notes. The GenBank sequence DQ283258 View Materials with the voucher identified as Cacosternum platys (Frost et al. 2006) is Microbatrachella capensis . The GenBank sequence DQ 022352 View Materials with the voucher identified as Cacosternum boettgeri (Scott 2005) is C. platys .
Genetics. This species varies from the other 14 species for the 16S fragment, by uncorrected p values of 2.5–5.9%. A single specimen had only one likely tyr haplotype, that was not shared with any other species.
Advertisement call. A series of 29 calls of four individuals from the Serengeti National Park, Tanzania showed a mean duration of 0.55 s (range 0.37–0.77) and the mean number of notes (pulse trains) 6.2 (range 4–8) (Channing et al. 2005). A typical advertisement call from Nairobi ( Fig. 11 View FIGURE 11 ) is a harsh clicking, where each series of notes consists of double pulses, apart from the initial note of three pulses. The call duration is 0.5 s, with six notes at a rate of 10 s -1.
Morphology. The female holotype has a SUL 19.7 (Channing et al. 2005). Breeding males have yellow vocal sacs without a pale margin ( Fig. 4 View FIGURE 4 ). The dorsum is smooth, and, viewed from above the nostrils are positioned on the anterior edge of the snout. The supratympanic fold is inconspicuous. The rictal gland is indistinct. There are 5 supernumerary tubercles on the palm. There is no webbing, with the notch between the third and fourth toes reaching the proximal subarticular tubercle of the third toe. The outer metatarsal is absent or indistinct, while the inner metatarsal tubercle is conical and protruding, twice the width of the tip of the first toe. The nuptial pad in breeding males is slightly pigmented but massive, reaching the proximal subarticular tubercle. The belly markings consist of groupings of brown speckles that form blotches with spots equal to the size of the eye lens.
Distribution. The species appears to be present from northern Tanzania to the highlands of central Kenya (Channing et al. 2005, Channing & Schmitz 2009) ( Fig. 6 View FIGURE 6 ) and may be more widely distributed. It breeds in temporary and permanent pools with little emergent vegetation.
Cacosternum rhythmum sp. nov. Rhythmic Dainty Frog. ( Figs. 3 View FIGURE 3 N, 4N)
Holotype. An adult male NMB A7521, collected by M. Burger, 18 December 2001 at Nottingham Road (29° 21’ 21” S, 29° 59’ 36” E), KwaZulu-Natal Province, South Africa. The habitat is flat open terrain with shallow pools and flooded grassy patches.
Paratypes. Eight males and one female, NMB A7522–7530, from Nottingham Road, Nottingham Ford, and 28 km S of Harrismith.
Diagnosis. This species differs from the other 14 species for the 16S fragment, by uncorrected p values of 1.1– 6.2%. The within-clade variation ranges from 0.0–0.2%. The small difference of 1.1% with C. boettgeri will be considered in the Discussion. The sample had eight likely tyr haplotypes, of which two were shared between one C. nanum and one C. rhythmum sp. nov. The vocal sac of breeding males is white ( Fig. 5 View FIGURE 5 ), which distinguishes it from species where the sac is black, grey or yellowish-beige ( C. capense , C. karooicum , C. leleupi , C. namaquense , C. nanogularum sp. nov., C. aggestum sp. nov., C. boettgeri , C. australis sp nov., C. nanum , C. parvum , C. platys , and C. kinangopensis . It is distinguished from C. capense by the absence of protruding dorsolateral glands and a pair just above the vent. Viewed from above, the nostrils are close to the anterior margin of the snout, distinguishing it from C. karooicum , which has the nostrils about three nostril diameters back from the anterior edge of the snout. The supratympanic fold continues posteriorly as a weak saddle, distinguishing it from C. nanogularum sp. nov., which has no saddle. The rictal gland is smooth, tapering to the arm insertion. This distinguishes it from those species where the rictal gland is interrupted ( C. capense , C. karooicum , C. striatum , C. nanum , C. parvum and C.platys ). There are no supernumerary tubercles on the palm, distinguishing it from species with 2–5 tubercles ( C. capense , C. karooicum , C. leleupi , C. namaquense , C. nanogularum sp. nov., C. aggestum sp. nov., C. boettgeri , C. nanum and C. platys ). It has no webbing, which distinguishes it from C. leleupi , which has a trace of webbing between the third and fourth toes. The inner metatarsal tubercle is small, conical, equal in width to the proximal subarticular tubercle of the first toe. This distinguishes it from species where the width of the inner metatarsal tubercle is equal to the width of the tip of the first toe ( C. capense , C. karooicum , C. namaquense , C. nanogularum sp. nov., C. australis sp. nov., C. striatum , C. nanum , C. plimptoni and C. platys ), and from those where the inner metatarsal tubercle is twice the width of the tip of the first toe ( C. leleupi , C. aggestum sp. nov., and C. parvum . The belly is pale, with a few very small brown blotches. This distinguishes it from species where the spots are larger than the eye lens, sometimes running together to form blotches ( C. capense , C. namaquense , C. nanogularum sp. nov., C. aggestum C. aggestum sp. nov.,) and from C. australis sp. nov. which has irregular yellow-beige blotches overlain with very small black speckles, and from C. striatum which has a silvery-white belly with small black blotches.
The advertisement call is complex and long. A typical call has a duration over 1.5 s, and consists of a string of pulses followed by a number of chirps, produced with a distinctive beat. It differs from the simple series of single or multiple clicks of C. boettgeri , C. kinangopensis , C. leleupi , and C. plimptoni or the creaking calls of C. capense , C. karooicum , C. namaquense , C. nanogularum sp. nov., C. parvum , and C. striatum or the brief chirp of C. nanum . It differs from those species producing a 'bouncing marble' call of pulses that speed up such as C. aggestum sp. nov., C. australis sp. nov. and C. platys .
Description of the holotype. The holotype is an adult male, SUL 15.0. Body gracile, widest at mid-belly, with a narrow head (HW/SUL 0.34). The head is acutely rounded from above and in profile. Head length measured from the angle of the jaw is moderate (HL/SUL 0.32). Canthus rostralis rounded, straight from eye to nostril, loreal region sloped outwards; nostrils small, rounded, directed laterally. The nostrils are placed closer to the snout than the eye (EN/SL 0.61). Internostril distance is less than distance between eye and nostril (NN/EN 0.91). Eyes directed anterolaterally, the eyes protrude, and are visible from below, relatively small (ED/HW 0.35; ED/SUL 0.12), equal to snout (ED/SL 1.0). Distance between anterior corners of eyes greater than internostril distance (NN/ EE 0.36). The angle of the jaw is posterior to a line drawn vertically from the back of the eye. The tympanum is not visible. Jaws without dentition; choanae small, round, located at anterior margins of roof of mouth; vomer processes and teeth absent; tongue long, narrow, slightly bifurcated distally. No median lingual papilla present.
The dorsal surfaces of the head, trunk and limbs are smooth, with glands and skin folds present; the rictal gland is smooth, continuing posteriorly unbroken to the arm insertion. The supratympanic fold forms a groove running over the arm to the leg insertion. Lines of broken skin glands form an hourglass pattern dorsally, narrowest at the scapular region, with a second pair of broken skin folds parallel to the midline, running from behind the scapular region nearly to the tip of the urostyle. The underside is smooth, with a slightly pigmented vocal sac with lateral folds ( Fig. 4 View FIGURE 4 ).
The forelimb is slender, hand small (HAN/SUL 0.26), finger tips bluntly rounded without discs. Relative finger lengths I<II<IV<III; subarticular tubercles distinct, rounded, with one on fingers I and II, two on finger IV and three on finger III, with the proximal subarticular tubercle on finger III small but distinct. No webbing between fingers. Thenar tubercle small, rounded, partially obscured by nuptial pad that reaches the distal phalanx of the first finger; palmar tubercles and inner metatarsal tubercle small, rounded, outer metacarpal tubercle absent. There are no supernumerary tubercles on the palm. An outgrowth is present on the outside of the fourth finger, at the level of the penultimate phalanx. It resembles a subarticular tubercle. No other growths like this are present in the sample examined.
Hind limbs moderately long (TIB/SUL 0.43; FOT/SUL 0.53), foot longer than tibia (TIB/FOT 0.82); thighs are moderately developed, with rough glands on the inner posterior faces; relative toe lengths are I<II<V<III<IV. The toe tips are not expanded; subarticular tubercles: one on toes I and II, two on toes III and V, and three on toe IV. Only a trace of webbing between the toes. Inner metatarsal tubercle conical, prominent, outer metatarsal tubercle present as a small pale raised spot.
Colour in preservative. The back has a grey background, with a darker brown vertebral band, within which is a thin pale vertebral line. Small darker patches are arranged in pairs. The belly is immaculate, with a margin of small clouds of speckles arranged in spots. The undersides of the hands and feet are pigmented. Colour in life. The dorsum is tan and brown with small black speckles and white blotches.
Paratype variation. The paratypes are similar in body proportions to the holotype. The males range in SUL from 15.3–16.7, with the female 19.3. The rictal gland is identical in all the paratypes, while the subarticular tubercles of the hand vary slightly, from round to a sharper cone-shape. The female has a uniform coloured back, but all the male paratypes have a wide vertebral band, or a narrower vertebral line. Two males have dorsal warts. The belly is immaculate in eight paratypes, with one possessing a few small spots, leaving over 90% of the belly unpigmented. One specimen, although immaculate, possesses light clouds of speckles shaped as spots. The specimens show the darker belly window through which the abdominal muscles can be seen, and all have supernumerary tubercles on the hands. Eggs are visible through the belly skin of the female.
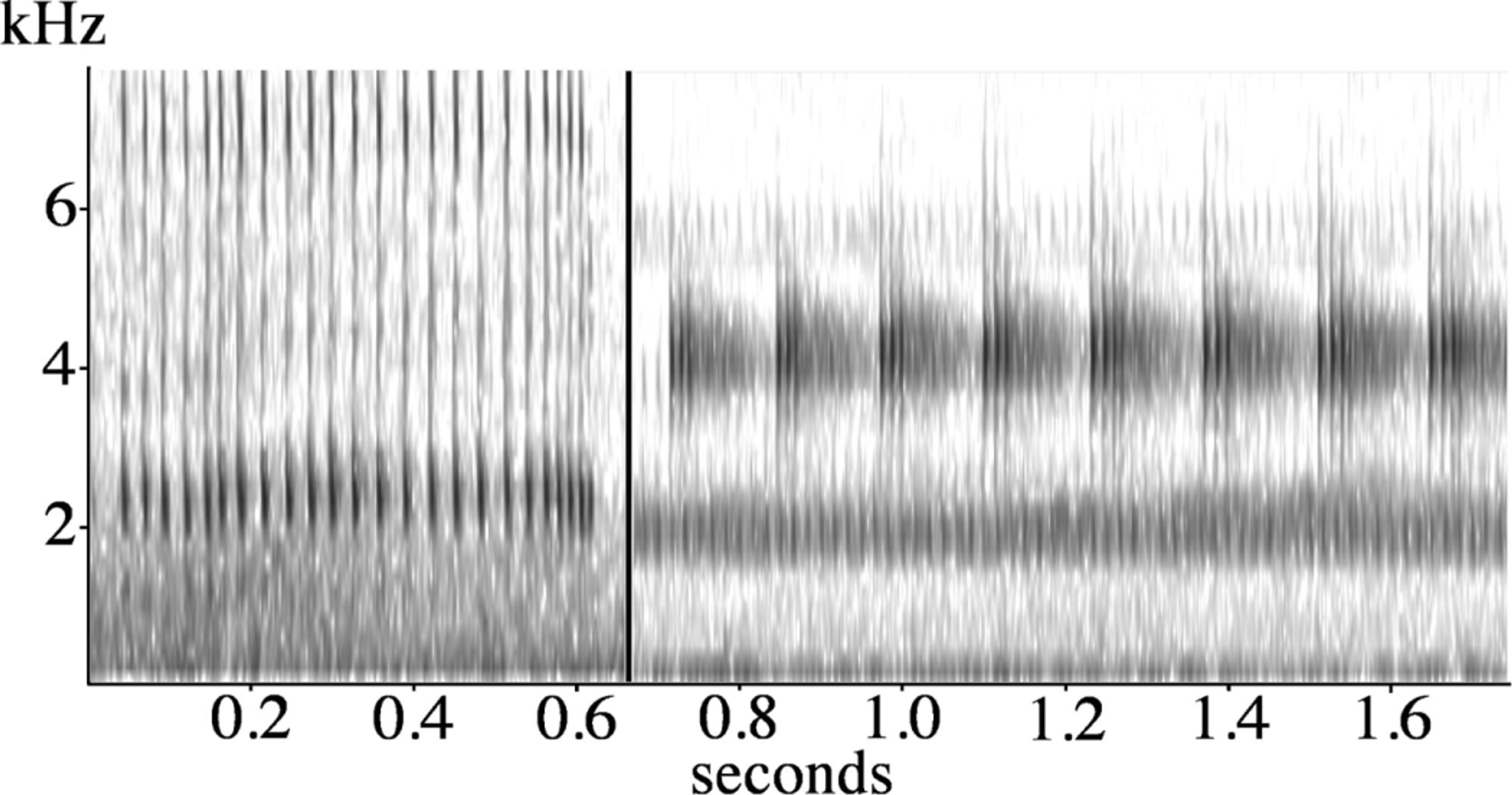
Advertisement call. A typical call recorded at Nottingham Road is illustrated in Fig 12 View FIGURE 12 . The call consists of an initial phase of single or double clicks lasting about one second, at a rate of 10 notes s -1, followed by three chirps, each with seven pulses. The pulse rate of each chirp is about 60 s -1, with the chirps uttered at a rate of 6 s -1. The duration of such a call is 1.6 s. The call has a distinct repeated beat, characteristic of this species.
Eggs and tadpoles. Unknown.
Distribution. Our specimens come from the KwaZulu-Natal midlands in South Africa ( Fig. 7 View FIGURE 7 ), but we expect the species to be more widely distributed. It breeds in temporary pools with abundant vegetation.
Etymology. The species name is derived from the Latin rhythmus (rhythm) referring to the rhythmic advertisement call. It is a noun in apposition.
Note. A call description for Cacosternum cf. striatum from Champagne Castle (Pickersgill 2007) is attributable to C. rhythmum sp. nov.








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.