
Solemya (Petrasma) cf. woodwardiana Leckenby, 1859
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3859.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:24FCAAE1-AB7C-4FAD-8698-D0C9F12400EC |
|
DOI |
https://doi.org/10.5281/zenodo.5228239 |
|
persistent identifier |
https://treatment.plazi.org/id/A2311D4D-9F1E-E32E-04E7-FB2DFCA3289A |
|
treatment provided by |
Felipe |
|
scientific name |
Solemya (Petrasma) cf. woodwardiana Leckenby, 1859 |
| status |
|
Solemya (Petrasma) cf. woodwardiana Leckenby, 1859
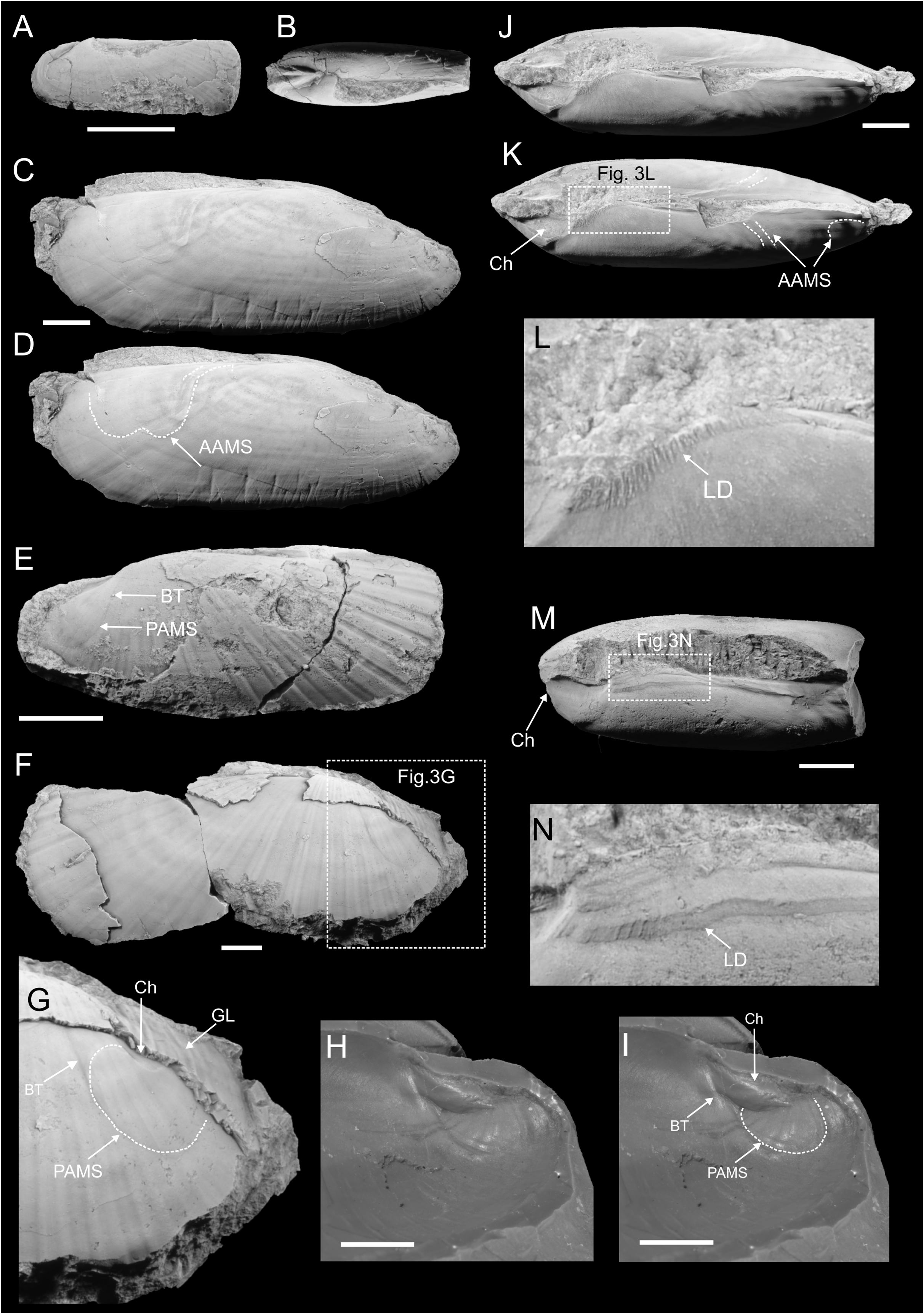
( Figure 3 View FIGURE 3 )
1859 cf. Solemya woodwardiana sp. nov. —Leckenby, p. 14, pl. 3, fig. 7.
1978 cf. Solemya woodwardiana Leckenby —Duff, p. 31, pl. 1, figs. 34–44.
? 1990 Solemya cf. woodwardiana Leckenby —Wignall, p. 8.
? 1993 Solemya cf. woodwardiana Leckenby —Wignall & Pickering, p. 328.
2011 Solemyid—Hammer et al., fig. 7h, tab. 2.
Material examined. 26 specimens, articulated to partially articulated; mostly internal moulds with only small portions of shell preserved. See Appendix 1 for the list of specimens.
Dimensions. 14–62 mm in length, 4.2–32 mm in width, 4–23.5 mm in height. See Figure 4 A–C View FIGURE 4 and Appendix 2A for details.
Description. Shell medium to large, elongated and narrow. Anterior margin broadly rounded, with anteriormost extremity positioned roughly in middle of curvature. Dorsal and ventral margin parallel to subparallel. Valves of some partially articulated specimens rotated along ligament line, which gives a false impression of shell widening anteriorly. Posterior margin evenly rounded, with curvature more pronounced than that of anterior margin. Umbones weak, opisthogyrate. External ornament composed of radial ridges projecting from umbonal area towards shell margins and commarginal growth lines, preserved as weak and broad wrinkles. Radial ridges are two to three times as wide as interridge spaces. On internal moulds external radial ornament is weaker. Anterior adductor muscle scar weak and positioned close to anterior shell extremity, supported by dorsal margin. Anterior adductor muscle scar rarely preserved well enough to define shape, but when present forms a broad arch pointing anteriorly, with its posteroventral margin running dorsally, passing into oblique and elongated portion, probably representing a visceral mass integument attachment scar. Posterior adductor muscle scar well impressed, especially on larger specimens, egg-shaped, with acute end pointing towards umbo. Anterior margin of posterior adductor muscle scar supported by well-developed buttress, running obliquely in dorsal direction and connected with chondrophore. Dorsal margin of posterior adductor muscle scar supported on a chondrophore; posterodorsal margin of posterior adductor muscle scar projecting above chondrophore. Hinge edentulous. Ligament internal, supported on a chondrophore, with small ligament demipads parallel to dorsal margin anterior of chondrophore.
Remarks. We compare our species with Solemya woodwardiana Leckenby, 1859 , from the Callovian of England, due to the overall similar shape and shell proportions. Duff (1978) noted the presence of a chondrophore and a buttress running along the anterior edge of posterior adductor muscle scar in S. woodwardiana . However, we leave our material in open nomenclature, because not all important solemyid characters are sufficiently defined in the Callovian S. woodwardiana . A similar species, identified as S. cf. woodwardiana , comes from the Kimmeridgian organic-rich facies of England ( Wignall 1990) and Scotland ( Wignall & Pickering 1993). Solemya species with shapes similar to S. woodwardiana are common in the Boreal Jurassic and Cretaceous ( Figure 4A View FIGURE 4 ). Unio togata from the Moscow area in Russia ( Trautschold 1858, p. 550, pl. 4, fig. 3) is a solemyid, with a characteristic notch in the subumbonal area, possibly indicating a presence of a buttress in front of PAMS. Trautschold (1858), probably unaware of Lamarck’s (1818) designation of the genus Solemya , synonymized his finds with the recent solemyid Tellina togata Poli, 1795 = Solemya togata ( Poli, 1795) . Another solemyid with a shape comparable to the Svalbard seep specimens is Solemya strigata Lahusen, 1886 , from the Bajocian around the Olenek and Lena River mouths ( Lahusen 1886; Zakharov & Shurigin 1978). A solemyid with similar shape and proportions was also noted from the Volgian of East Greenland by Fürsich (1982). Solemya cf. woodwardiana differs from the Solemya voltzii ( Roemer, 1839) from the Toarcian Posidonienschiefer of Germany by having beaks positioned closer to the posterior than that species. A species described by Lindström (1865) as Solenomya torelli from the Jurassic of Svalbard is more elongated than most of the specimens of S. cf. woodwardiana , and Lindström’s brief diagnosis lacks any information about the ligament morphology. There is no certainty about the stratigraphic position of this species, since there is no Jurassic sediments cropping out in the Adventfjorden [Advent Bay] area of Svalbard, where Lindström’s specimens are said to come from.
The external ornament of modern solemyids, especially of the thin-shelled species, is likely to be subjected to early diagenetic modification due to shell dissolution. The external shell shape and proportions can also be subjected to some variation due to plasticity and deformation of the organic-rich shell during burrowing (e.g. Stanley 1970; Taylor & Glover 2010), and by later compaction. Therefore, ascribing a bivalve fossil to the solemyids based on elongated shape only can be problematic. For example, Solenomya (?) hoeli from the Upper Jurassic of Svalbard ( Sokolov & Bodylevsky 1931; Birkenmajer et al. 1982) has a very similar shape and proportions to our Solemya cf. woodwardiana specimens, and also has a radial ornament. However, detailed investigation of Sokolov & Bodylevsky’s type specimen reveals that the radial pattern on the anterior part of the shell is in fact caused by cracking. Sokolov & Bodylevsky (1931) failed to notive very a fine radial ornament on the anterior and posterior margins of their type, which shows that Solenomya (?) hoeli is not a solemyid and instead belongs to the genus Musculus Roeding, 1798 .
Occurrence. Solemya woodwardiana: Callovian of England ( Leckenby 1859; Duff 1978). Solemya cf. woodwardiana: Kimmeridgian of England ( Wignall 1990) and Scotland ( Wignall & Pickering 1993); seeps 1, 3, 5, 8, 9 and 12 (Upper Volgian–uppermost Ryazanian), Slottsmøya Member, Svalbard ( Tab. 1 View TABLE 1 ).
Palaeoecology. We assume that the Svalbard seep Solemya cf. woodwardiana was chemosymbiotic, as are all known modern members of the family. Since species similar to Solemya cf. woodwardiana are present in organicrich, offshore facies of the Middle and Upper Jurassic of Northern Europe and the Arctic ( Trautschold 1858; Lahusen 1886; Duff 1978; Zakharov & Shurigin 1978; Wignall 1990; Wignall & Pickering 1993), we infer that Svalbard species did not have any particular affiliation with seep environments and had a broad ecological tolerance, populating high redox potential environments.
Solemyids are widespread in modern oceans from intertidal to abyssal depths ( Taylor & Glover 2010). The genus Solemya s.l. is a member of shelf and slope faunas ( Coan et al. 2000; Coan & Scott 2012), where it is present in soft substrates enriched in organic matter, like reduced sediments (e.g. Conway et al. 1992), wood debris accumulations ( Reid 1980), sea grass beds (e.g. Taylor et al. 2008) and carrion falls ( Fujiwara et al. 2009). The deepest confirmed record of Solemya s.l. comes from 1510 m in Sagami Bay ( Kamenev 2009), and it possibly occurs as deep as 1697 m in the Eastern Mediterranean ( Rodrigues et al. 2011).
Solemya s.l. is a burrower, especially favouring muddy, silty and sandy bottoms. Burrowing takes place anteriorfirst and leads to formation of U-to Y-shaped tubes, reinforced with mucus ( Stanley 1970; Reid 1980; Stewart & Cavanaugh 2006). While resting in its burrow, the animal uses its foot in a pumping action, supplying water enriched in reduced compounds from the lower shafts of the burrow to their gills ( Reid 1980). All investigated Solemya s.l. species live in symbiosis with thiotrophic Gammaproteobacteria ( Stewart & Cavanaugh 2006; Taylor & Glover 2010). A gutless condition has been confirmed in some species (e.g. Reid 1980; Conway et al. 1992; Kamenev 2009), while others retain the gut (e.g. Taylor et al. 2008; Oliver et al. 2011) and have limited ability for particulate feeding ( Krueger et al. 1992). Coan et al. (2000) noticed that gutless species of Solemya s.l. are more common in environments with extreme organic content.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |