
Arthromelodes tokushigei, Jałoszyński, 2023
|
publication ID |
https://doi.org/10.11646/zootaxa.5325.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:7C6BF79E-BE56-4A46-8746-CDA856CA8BF9 |
|
DOI |
https://doi.org/10.5281/zenodo.8247117 |
|
persistent identifier |
https://treatment.plazi.org/id/A02A8786-FFCC-DD64-FF51-78382D4BFAC6 |
|
treatment provided by |
Plazi |
|
scientific name |
Arthromelodes tokushigei |
| status |
sp. nov. |
Arthromelodes tokushigei sp. n.
( Figs 1–48 View FIGURES 1–2 View FIGURES 3–6 View FIGURES 7–10 View FIGURES 11–14 View FIGURES 15–17 View FIGURES 18–20 View FIGURES 21–26 View FIGURES 27–30 View FIGURES 31–36 View FIGURES 37–42 View FIGURES 43–48 )
Type material. Holotype: JAPAN ( Okinawa Pref.): ♁, two labels: “ JAPAN, OKINAWA Pref. / OKINAWAjima, 26 ii 2023 / Ohkuni-rindo, Jashiki / 2645’15.7’’N/12814’44.8’’E / leg. P. JAŁOSZYŃSKI” [white, printed], “ ARTHROMELODES / tokushigei m. / P. Jałoszyński, 2023/ HOLOTYPUS” [red, printed] ( NSMT) . Paratypes (10 exx.): 7 ♁♁ (one disarticulated), 3 ♀♀, same data as for holotype (cPJ, NSMT) .
Diagnosis. Punctures on dorsum of head dense and distinct in anterior half, posteriorly becoming shallower and sparser; punctures on pronotal disc distinct and dense but shallow; in males each profemur with elongate transversely carinulate field in basal half of ventral surface and mesotibia with elongate ventral apical tooth or mucro, but all trochanters unmodified; abdominal tergite IV in male with large posterior transverse impression bearing large oval median tubercle near posterior margin of tergite and three pairs of minute elongate processes in front of tubercle, lateral setal patches of male tergite IV situated entirely laterally to posteromedian impression; aedeagus in ventral view with ventral projection of median lobe (inserted just above orifice) recurved and with subtriangular apical region with strongly oblique distal margin, dorsal projection in lateral view bent ventrad nearly at right angle and with minute subtriangular proximal process.
Description. Body of male ( Fig. 1 View FIGURES 1–2 ) elongate, strongly convex, moderately to darkly brown, with tarsi slightly lighter, setae slightly lighter than cuticle; BL 2.04–2.19 mm.
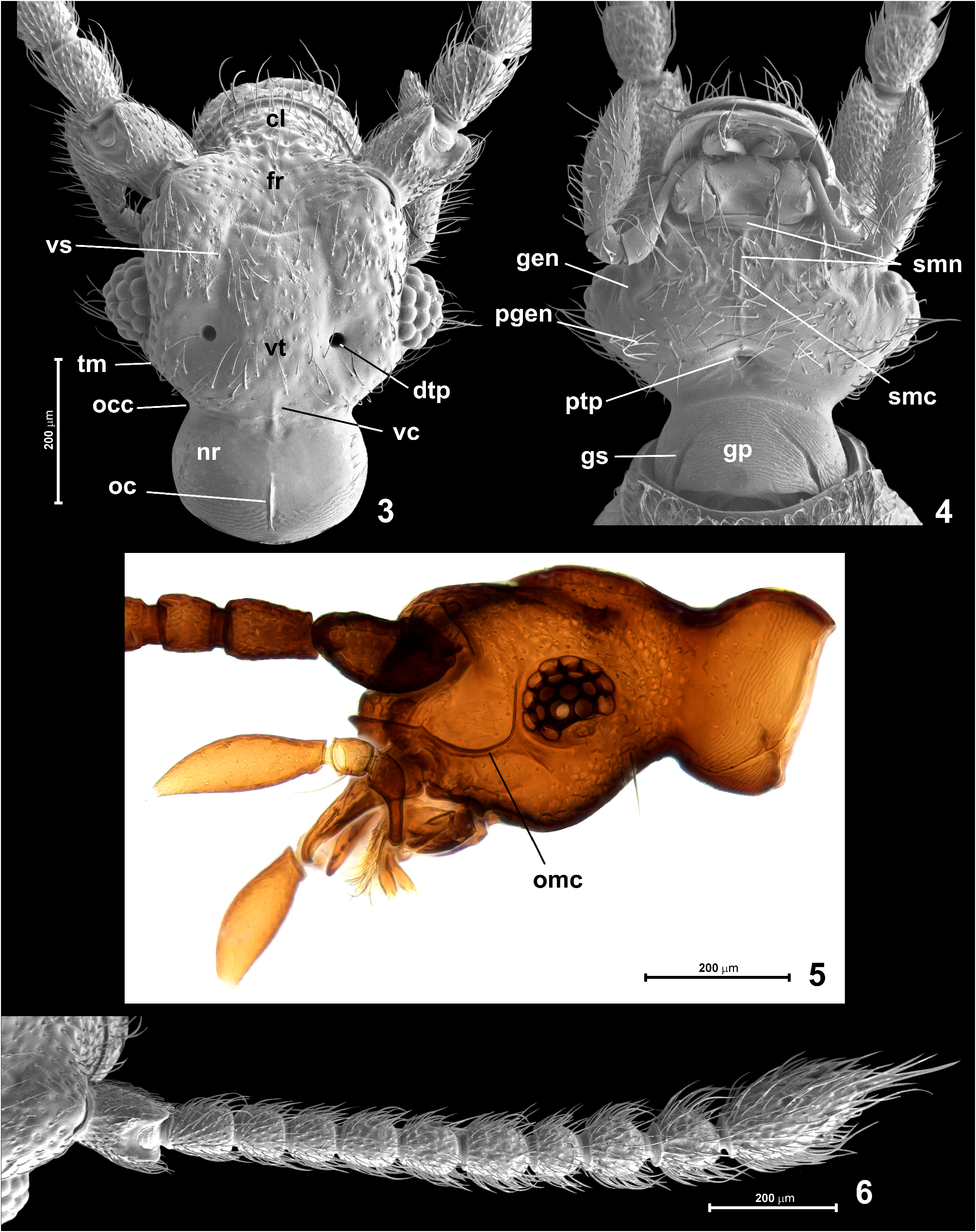
Head ( Figs 3–5 View FIGURES 3–6 ) subhexagonal, flattened, broadest at eyes, HL 0.41–0.48 mm, HW 0.44–0.45 mm. Neck region ( Fig. 3 View FIGURES 3–6 ; nr) dorsally smooth and with short posteromedian occipital carina ( Fig. 3 View FIGURES 3–6 ; occ); ventrally neck region with distinct gular sutures ( Fig. 4 View FIGURES 3–6 ; gs) obliterated anteriorly and demarcating subtriangular gular plate ( Fig. 4 View FIGURES 3–6 ; gp) covered with strongly transverse microreticulation; vertex ( Fig. 3 View FIGURES 3–6 ; vt) weakly convex medially and distinctly impressed laterally, with short median longitudinal vertexal carina ( Fig. 3 View FIGURES 3–6 ; vc) adjacent to posterior margin of vertex and not connected with occipital carina; dorsal tentorial pits ( Fig. 3 View FIGURES 3–6 ; dtp) large and deep, circular and asetose, each situated in lateral impressed region of vertex; vertex in front of dorsal tentorial pits and posterior margin of frons ( Fig. 3 View FIGURES 3–6 ; fr) in front of eyes with shallow vertexal sulcus ( Fig. 3 View FIGURES 3–6 ; vs) forming M-shaped figure with shallow transverse portion; frons at middle slightly impressed, impressed region flanked by prominent supraantennal tubercles. Eyes ( Figs 3–5 View FIGURES 3–6 ) large, strongly convex, bean-shaped, each with 18 convex corneal lenses. Occipital constriction ( Fig. 3 View FIGURES 3–6 ; occ) about as wide as 1/3 of HW including eyes; tempora ( Fig. 3 View FIGURES 3–6 ; tm) in dorsal view nearly as long as length of eye, nearly straight and strongly converging posterad. Clypeus ( Fig. 3 View FIGURES 3–6 ; cl) semicircular with rounded and distinctly carinate and upturned anterior margin. Punctures on dorsum of head dense and composed of large deep punctures on clypeus, posteriorly on frons and vertex gradually reducing in depth and becoming sparser, so that posterior margin of vertex remains virtually impunctate; setae on clypeus, frons and vertex sparse, short and suberect. Genae and postgenae ( Fig. 4 View FIGURES 3–6 ; gen, pgen) weakly convex and sparsely, distinctly punctate, sparsely covered with long and erect setae directed mainly laterally and anterolaterally. Antennal fossa conspicuously large and ventrally accentuated by distinct ocular-mandibular carina ( Fig. 5 View FIGURES 3–6 ; omc) running from ventral margin of eye to mandibular base and anteriorly confluent with marginal carina of clypeus. Posterior tentorial pits (marking border between gula and submentum) fused at middle to form single slightly transverse pit situated at level of posterior margins of eyes.
Antennae ( Figs 1 View FIGURES 1–2 , 6 View FIGURES 3–6 ) slightly shorter than head, pronotum and elytra together, weakly thickening toward apices; AnL 1.20–1.23 mm; scape distinctly elongate, subcylindrical, with deep dorsolateral emargination and flattening, pedicel slightly broadening distad, distinctly elongate but clearly shorter and narrower than scape; antennomeres 3–4 each about as long as broad, antennomeres 5–7 each slightly elongate (7 longest), antennomere 8 distinctly shorter than 7, about as long as broad, antennomeres 9–10 each about as long as broad, antennomere 11 nearly as long as 9–10 combined, distinctly broader than 10, fusiform, unmodified. All antennomeres covered with coarse microreticulation and dense setae, those on antennomeres 9–11 longer than those on proximal antennomeres.
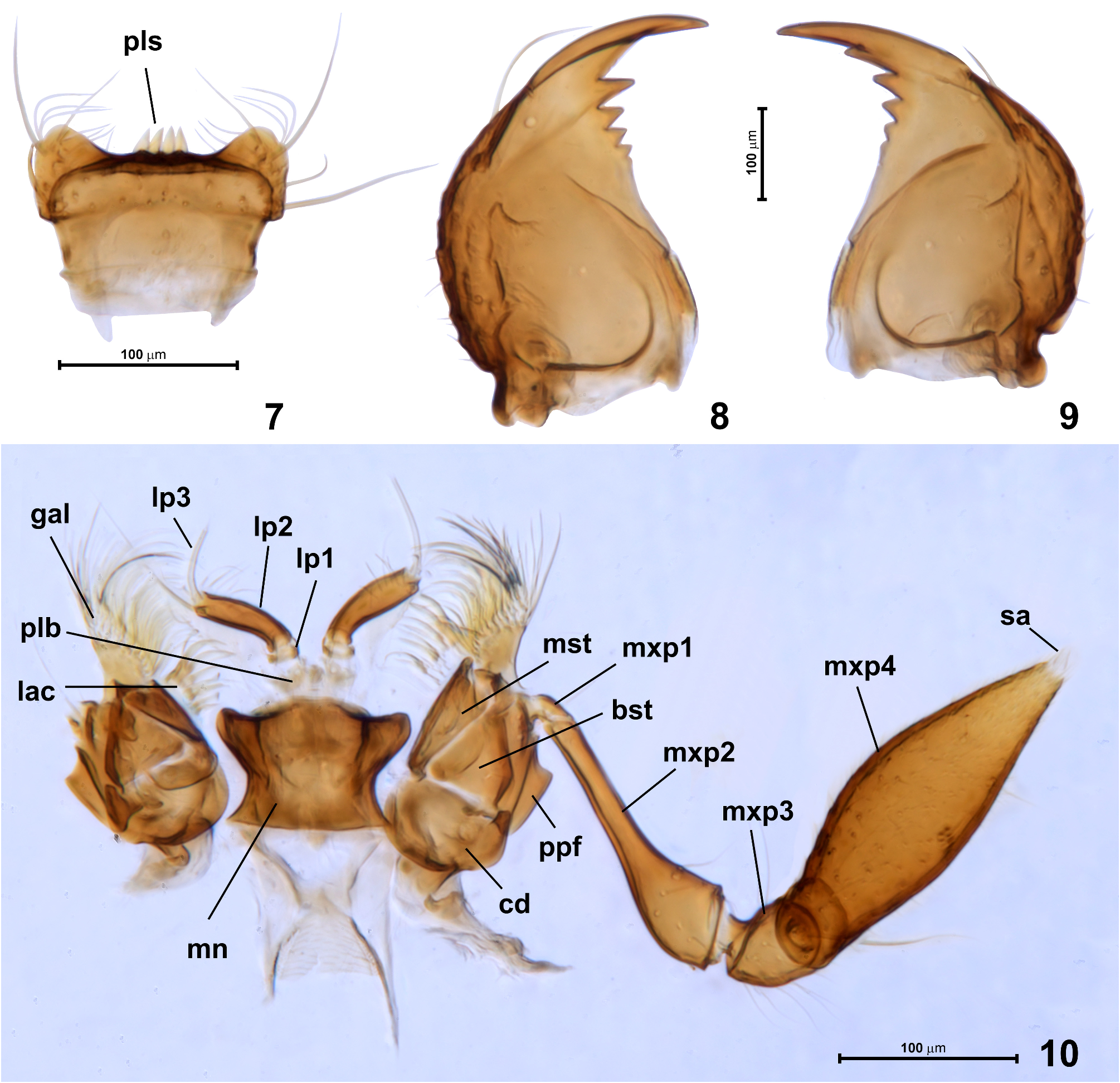
Labrum ( Fig. 7 View FIGURES 7–10 ) steeply declining anterad, broadening anteriorly, strongly transverse, with distinct subtriangular anterolateral lobes and nearly straight anterior margin; dorsal surface with sparse paired setae, four peg-like sensilla ( Fig. 7 View FIGURES 7–10 ; pls) of anterior epipharynx long, apically pointed and adjacent to each other.
Mandibles ( Figs 8, 9 View FIGURES 7–10 ) subtriangular, flattened dorsoventrally, each with sharp apical incisor and preapical row of four teeth reducing in size toward mandibular base; outer mandibular margin strongly convex and indistinctly undulate, with long seta inserted dorsolaterally in distal 1/3 of mandible, and a few short setae distributed along outer margin in posterior half.
Each maxilla( Fig.10 View FIGURES 7–10 )with transverse cardo( Fig.10 View FIGURES 7–10 ; cd),subtriangular basistipes( Fig.10 View FIGURES 7–10 ; bst)and subrectangular, elongate mediostipes (Fog. 10; mst); palpifer ( Fig. 10 View FIGURES 7–10 ; ppf) elongate, clearly demarcated from stipes; galea ( Fig. 10 View FIGURES 7–10 ; gal) weakly elongate, with dense fringe of long setae along distal and mesal margins; lacinia ( Fig. 10 View FIGURES 7–10 ; lac) much shorter than galea and with sparser and thicker setae along mesal margin. Maxillary palp ( Figs 4 View FIGURES 3–6 , 10 View FIGURES 7–10 ) slightly longer than exposed anterior region of head capsule; palpomere 1 ( Fig. 10 View FIGURES 7–10 ; mxp1) minute, elongate and asetose; palpomere 2 ( Fig. 10 View FIGURES 7–10 ; mxp2) pipe-shaped, strongly elongate and weakly curved in distal, broadened region, with distal margin oblique and carinate, surface covered with several sparsely distributed setae; palpomere 3 ( Fig. 10 View FIGURES 7–10 ; mxp3) small, subtriangular in lateral view, with sparse long setae distributed mainly on posterior margin; palpomere 4 ( Fig. 10 View FIGURES 7–10 ; mxp4) fusiform, strongly enlarged, covered with sparse and almost evenly distributed short setae and with 2–3 longer and more erect setae (in Fig. 10 View FIGURES 7–10 one long seta is visible near base); apical sensory appendage of palpomere 4 ( Fig. 10 View FIGURES 7–10 ; sa) minute, subconical.
Labium ( Figs 4 View FIGURES 3–6 , 10 View FIGURES 7–10 ) with submentum ( Fig. 4 View FIGURES 3–6 ; smn) discernible as short and slightly convex transverse stripe just behind mentum and prolonged posteriorly as narrow median submental carina ( Fig. 4 View FIGURES 3–6 ; smc) between strongly expanded mesally genae and postgenae; mentum ( Fig. 10 View FIGURES 7–10 ; mn) inversely subtrapezoidal with deep posterior constriction, so that posterior and anterior corners are projecting laterally, anterior margin convex at middle and emarginate at each side. Prelabium ( Fig. 10 View FIGURES 7–10 ; plb) largely membranous, with poorly discernible premental sclerites; insertions of labial palps narrowly separated; labial palps trimerous, palpomere 1 ( Fig. 10 View FIGURES 7–10 ; lp1) minute, annulate and asetose; palpomere 2 ( Fig. 10 View FIGURES 7–10 ; lp2) strongly elongate, weakly curved and slightly broadening towards apex, with several short and sparse setae mainly distributed in mesal apical region; palpomere 3 ( Fig. 10 View FIGURES 7–10 ; lp3) setiform, strongly elongate, slender, curved and asetose.
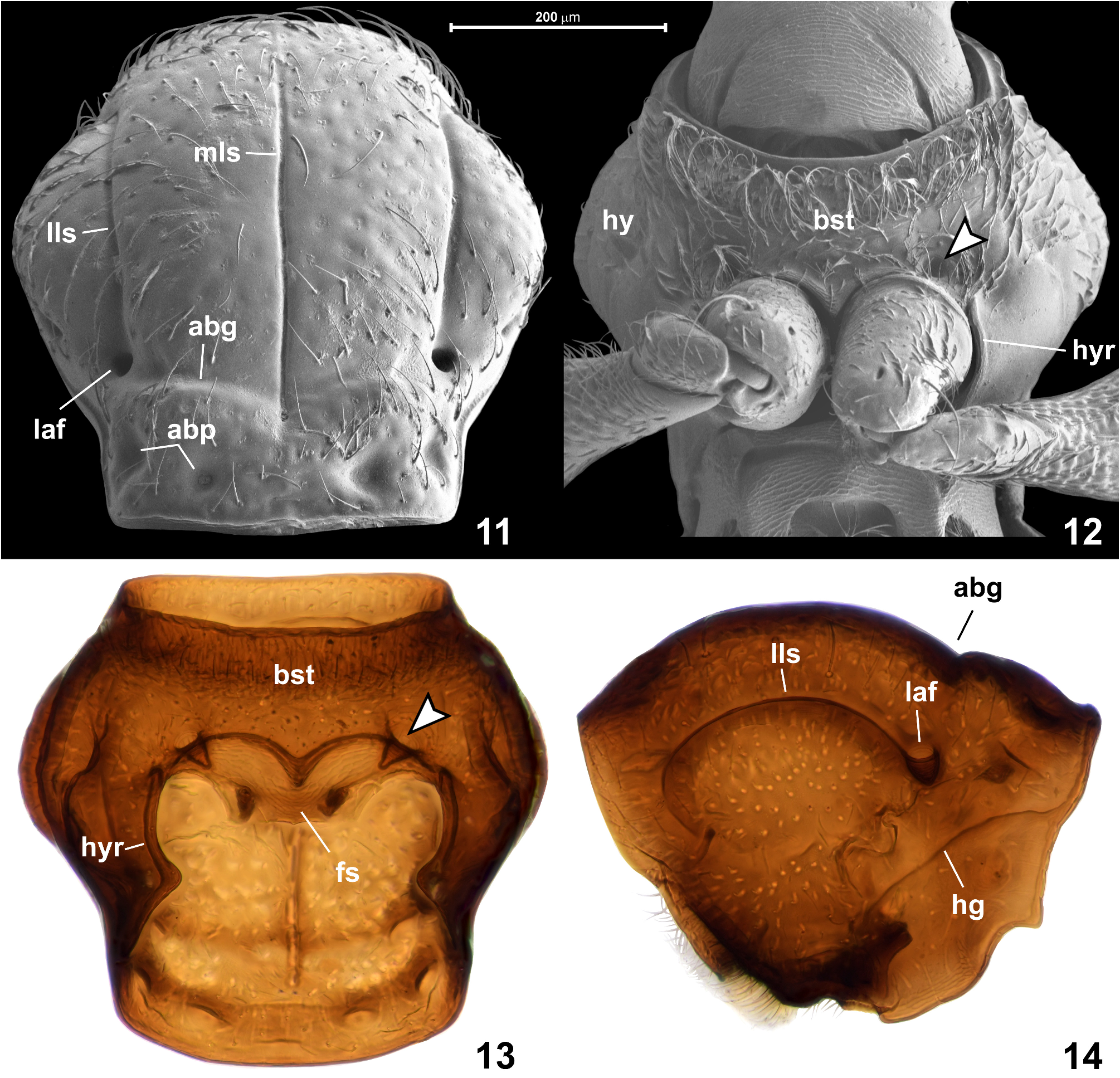
Prothorax ( Figs 11–14 View FIGURES 11–14 ) convex dorsally and flattened ventrally, slightly transverse, in dorsal view subhexagonal. Pronotum ( Figs 11, 14 View FIGURES 11–14 ) broadest distinctly in front of middle; PL 0.45–0.50 mm, PW 0.45–0.48 mm. Anterior margin nearly straight; lateral margins strongly convex in submedian region, slightly sinuate anteriorly and posteriorly; posterior corners nearly right-angled and blunt; posterior margin nearly straight, with marginal carina on entire length. Median longitudinal sulcus ( Fig. 11 View FIGURES 11–14 ; mls) narrow and sharply marked, anteriorly obliterated just before anterior pronotal margin and posteriorly deepened to form indistinct, vestigial and asetose median antebasal fovea; lateral longitudinal sulci ( Fig. 11 View FIGURES 11–14 ; lls) sharply marked and each posteriorly connected with large and asetose lateral antebasal fovea ( Figs 11, 14 View FIGURES 11–14 ; laf); lateral and median antebasal foveae connected by biarcuate and anteriorly convex antebasal transverse groove ( Fig. 11 View FIGURES 11–14 ; abg); pronotal base with inner and outer antebasal pits ( Fig. 11 View FIGURES 11–14 ; abp), each distinct and large but shallow and with diffuse margins. Punctures on disc distinct and large but shallow and somewhat diffuse, distances between punctures subequal to their diameters; setae sparse, short and suberect. Hypomera ( Fig. 12 View FIGURES 11–14 ; hy) anteriorly setose, posteriorly asetose.
Prosternum ( Figs 12, 13 View FIGURES 11–14 ) with basisternal (precoxal)region much longer than procoxal rests (which are composite structures combining posterior basisternal region and furcasternum situated behind profurcal invagination sites); notosternal sutures lacking; prosternum with one pair of foveae ( Figs 12, 13 View FIGURES 11–14 , indicated by arrowheads) situated just in front of and slightly mesad procoxal cavity, each directed posteromesad and with asetose opening. Basisternal region posteriorly forming short and broadly subtriangular prosternal process not separating procoxae; surface of basisternal region densely covered with long and recumbent to suberect setae, especially dense in anterior half of prosternum. Hypomeral ridge ( Figs 12, 13 View FIGURES 11–14 ; hyr) developed along adcoxal margin of each hypomeron and demarcating narrow inner region of hypomeron; hypomeral groove ( Fig. 14 View FIGURES 11–14 ; hg) present, but externally indistinct and poorly visible, in cleared specimens accentuated by line of darker cuticle marking internal strenghtening ridge.
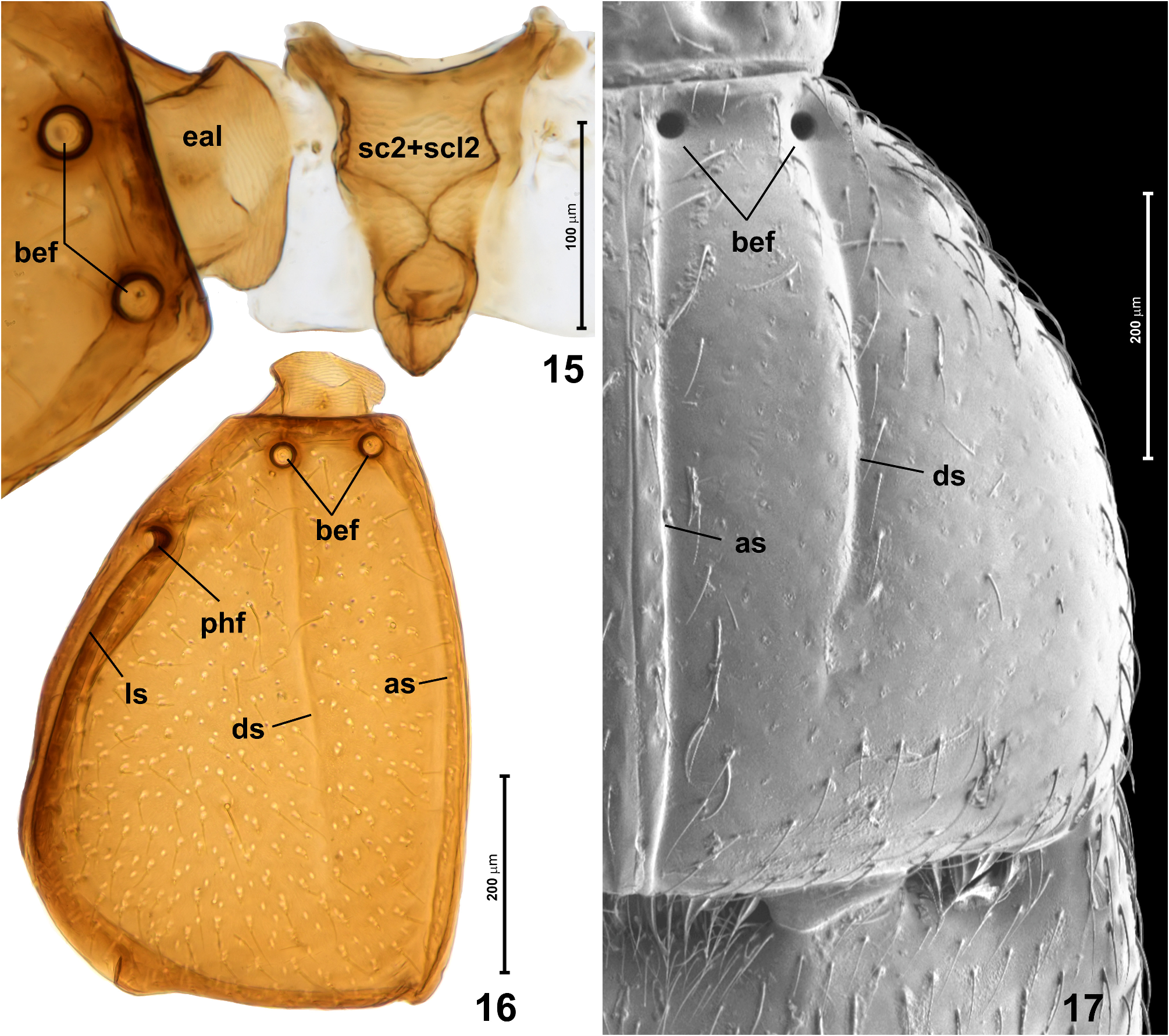
Mesonotum ( Fig. 15 View FIGURES 15–17 ) strongly elongate, subtriangular, lacking scutoscutellar suture; scutellar shield not exposed, hidden under posterior pronotal margin.
Elytra ( Figs 16, 17 View FIGURES 15–17 ) together transverse subtrapezoidal with strongly rounded sides, broadest near posterior third; EL 0.58–0.61 mm, EW 0.68–0.75 mm. Each elytron with complete adsutural sulcus ( Figs 16, 17 View FIGURES 15–17 ; as), discal sulcus (( Figs 16, 17 View FIGURES 15–17 ; ds) developed in anterior 2/3, lateral sulcus ( Fig. 16 View FIGURES 15–17 ; ls) not visible in dorsal view and extending from posthumeral fovea to posterolateral elytral corner. Each elytron with three distinct asetose circular foveae: pair of basal elytral foveae ( Figs 16, 17 View FIGURES 15–17 ; bef) situated in distinct transverse impression just behind anterior ridge of disc, and posthumeral fovea ( Fig. 16 View FIGURES 15–17 ; phf) situated on lateral elytral surface far behind humerus. Posterolateral elytral corner obtuse-angled and broadly rounded; posterior elytral margin weakly arcuate, with shallow emargination at adsutural and lateral corners. Punctures on elytra much less distinct than those on pronotal disc, superficial and inconspicuous; setae sparse, short and suberect.
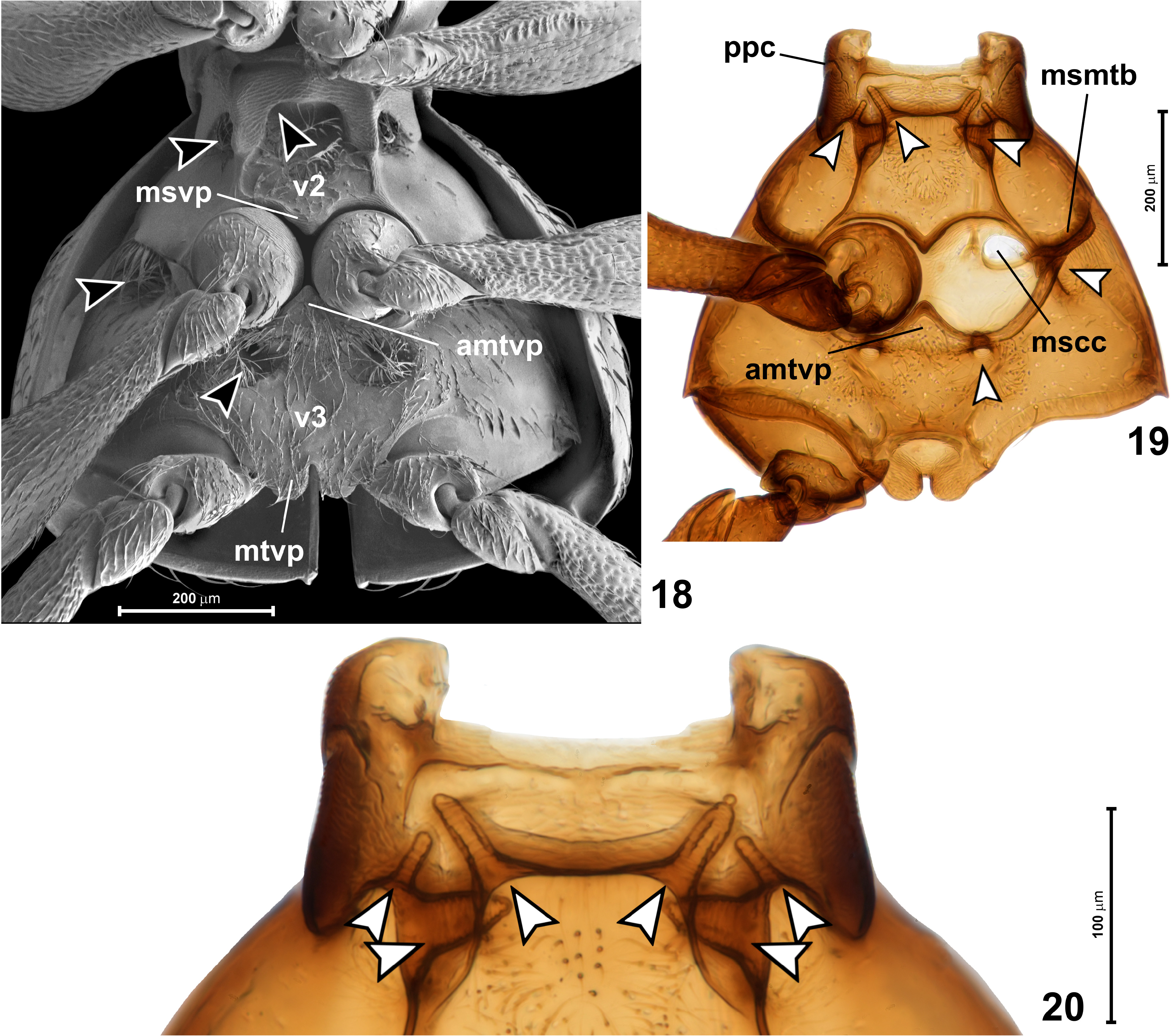
Mesoventrite ( Fig. 18 View FIGURES 18–20 ; v 2 View FIGURES 1–2 ) transverse, with lateral regions fully demarcated from metaventrite (i.e, mesometaventral border clearly marked; Fig. 19 View FIGURES 18–20 ; msmtb). Mesanepisterna and anterior region of mesoventrite forming massive prepectus ( Fig. 19 View FIGURES 18–20 ; ppc), posteriorly mesoventrite strongly and abruply broadeneing, with lateral margins strongly diverging posterad. Mesoventral process ( Fig. 18 View FIGURES 18–20 ; msvp) situated between anterior margins of mesocoxae, broadly subtriangular and posteriorly widely separated from anterior metaventral process. Prepectus covered with transverse microreticulation; lateral and posteromedian regions of mesoventrite smooth, sides asetose, median region covered with sparse suberect setae. Three pairs of foveae are situated on mesoventrite (indicated by arrowheads in Figs 18–20 View FIGURES 18–20 ): one pair ventrally and sublaterally just behind median transverse region of prepectus, directed anterolaterad; one pair ventrally and laterally between lateral region of prepectus and lateral asetose region of mesoventrite, directed anteromesad; and one pair sharing opening with the latter, but much broader and directed mesad. Opeinings of all mesoventral foveae densely setose.
Metanotum (not shown) strongly shortened and largely membranous, lacking alacristae and apodemes; hind wings entirely lacking.
Metaventrite ( Fig. 18 View FIGURES 18–20 ; v 3 View FIGURES 3–6 ) about twice as wide as long, weakly impressed at middle and convex at sides, with broadly subtriangular anterior metaventral (intermesocoxal) process ( Figs 18, 19 View FIGURES 18–20 ; amtvp). Metaventrite with two pairs of foveae (indicated by arrowheads in Figs 18, 19 View FIGURES 18–20 ): just behind and laterad mesocoxal cavities, directed masad and shallow, developed rather as deep impressions than typical foveal cuticular invaginations; and just behind middle of each mesocoxal rest, directed anterodorsad; openings of all metaventral foveae densely setose, those of posterior foveae situated in circular impressions much larger than diameter of fovea. Metaventral intermetacoxal process ( Fig. 18 View FIGURES 18–20 ; mtvp) broadly subtriangular, divided medially by narrow and long emargination into rounded lateral lobes. Metaventrite virtually impunctate, covered with sparse and short setae denser on median region than on sides.
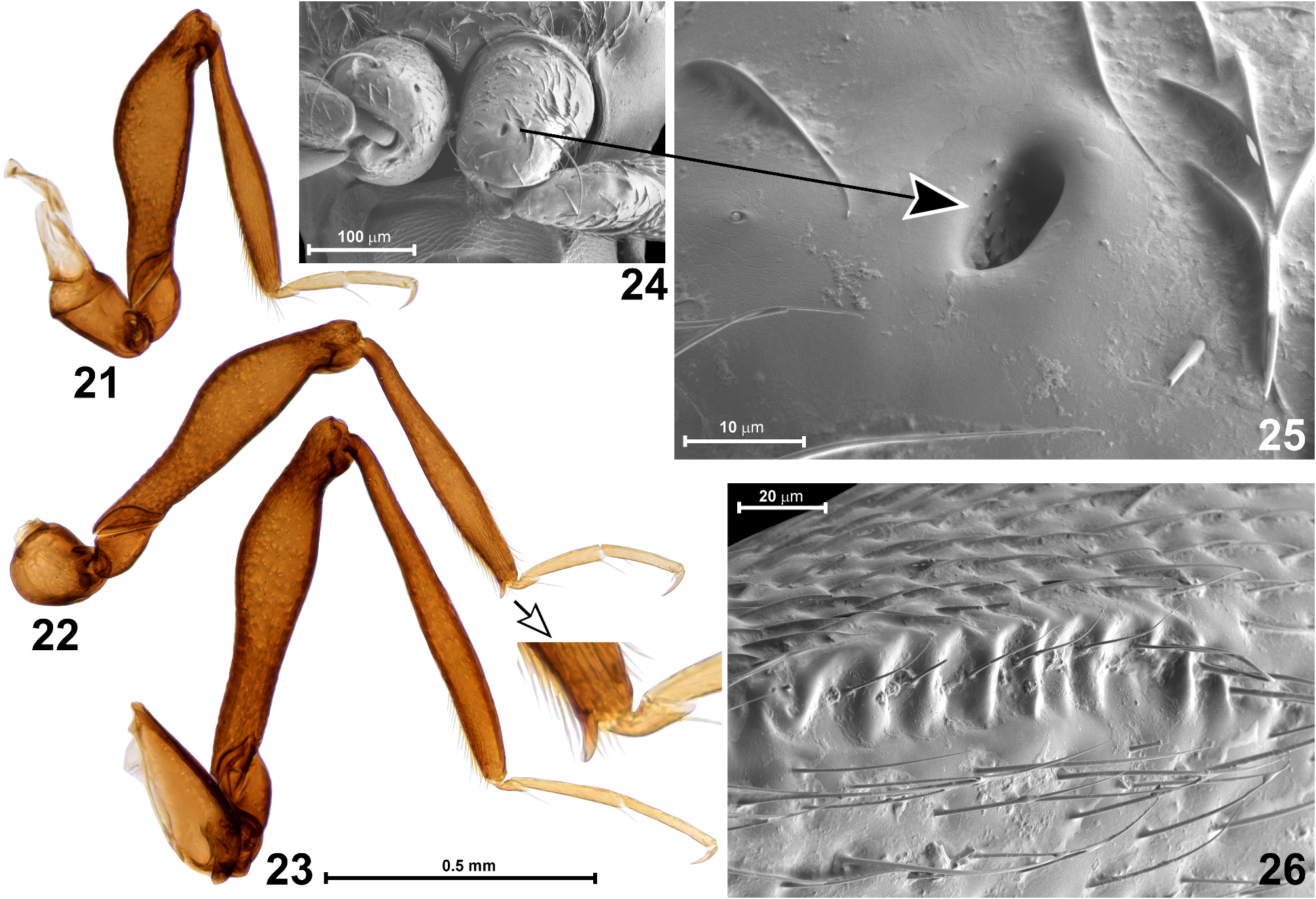
Legs ( Figs 21–26 View FIGURES 21–26 ) long and slender. Fore legs ( Fig. 21 View FIGURES 21–26 ) with subconical coxa proximally prolonged by trochantin fused with cryptopleuron (not visible in intact specimens), each procoxa with glandular or sensory asetose opening in distal region of anterior surface ( Figs 24, 25 View FIGURES 21–26 ); trochanter small, elongate subtriangular with rounded ventral margin; femur clavate, in proximal region of ventral surface with elongate field of transverse irregular carinulae ( Fig. 26 View FIGURES 21–26 ); tibia straight, broadening from base to around distal 1/3 and then tapering towards apex; tarsus slightly longer than half length of tibia, with tarsomere 3 indistinctly longer than 2. Middle leg ( Fig. 22 View FIGURES 21–26 ) similar to fore leg but femur ventrally smooth and tibia with slender ventral apical mucro. Hind leg ( Fig. 23 View FIGURES 21–26 ) longest and similar to fore and middle leg except for strongly transverse and subtriangular coxa and lack of any modifications.
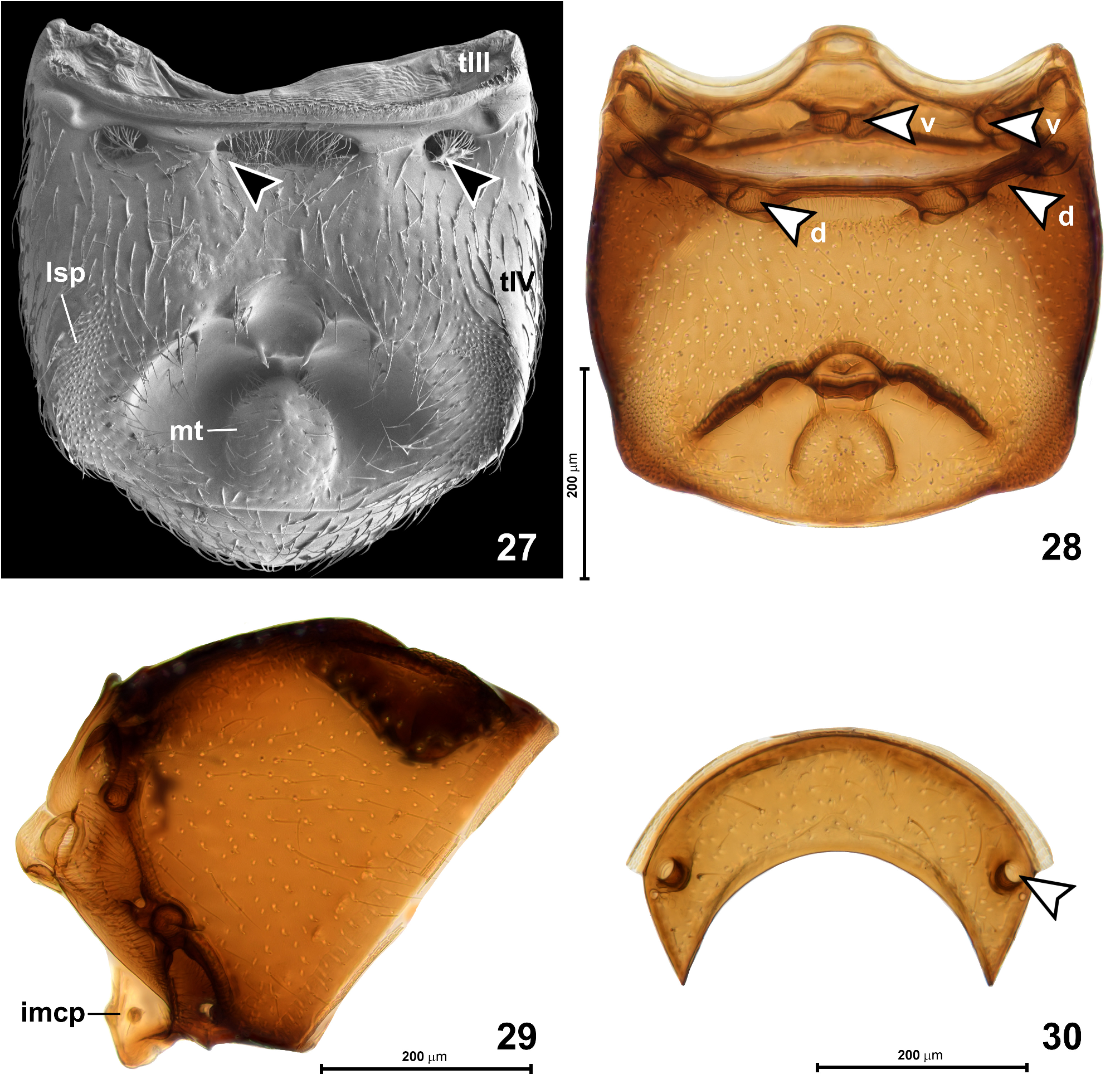
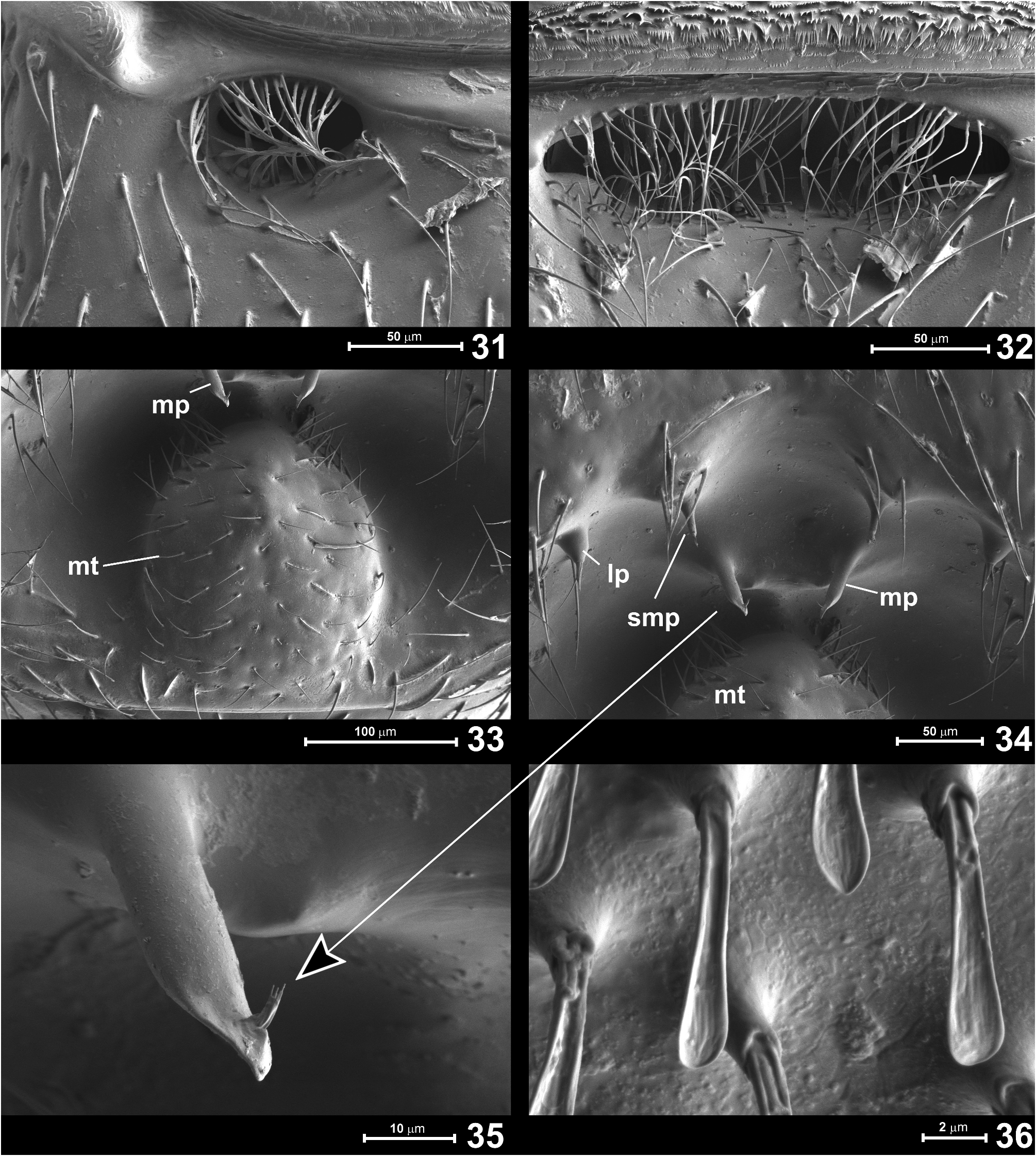
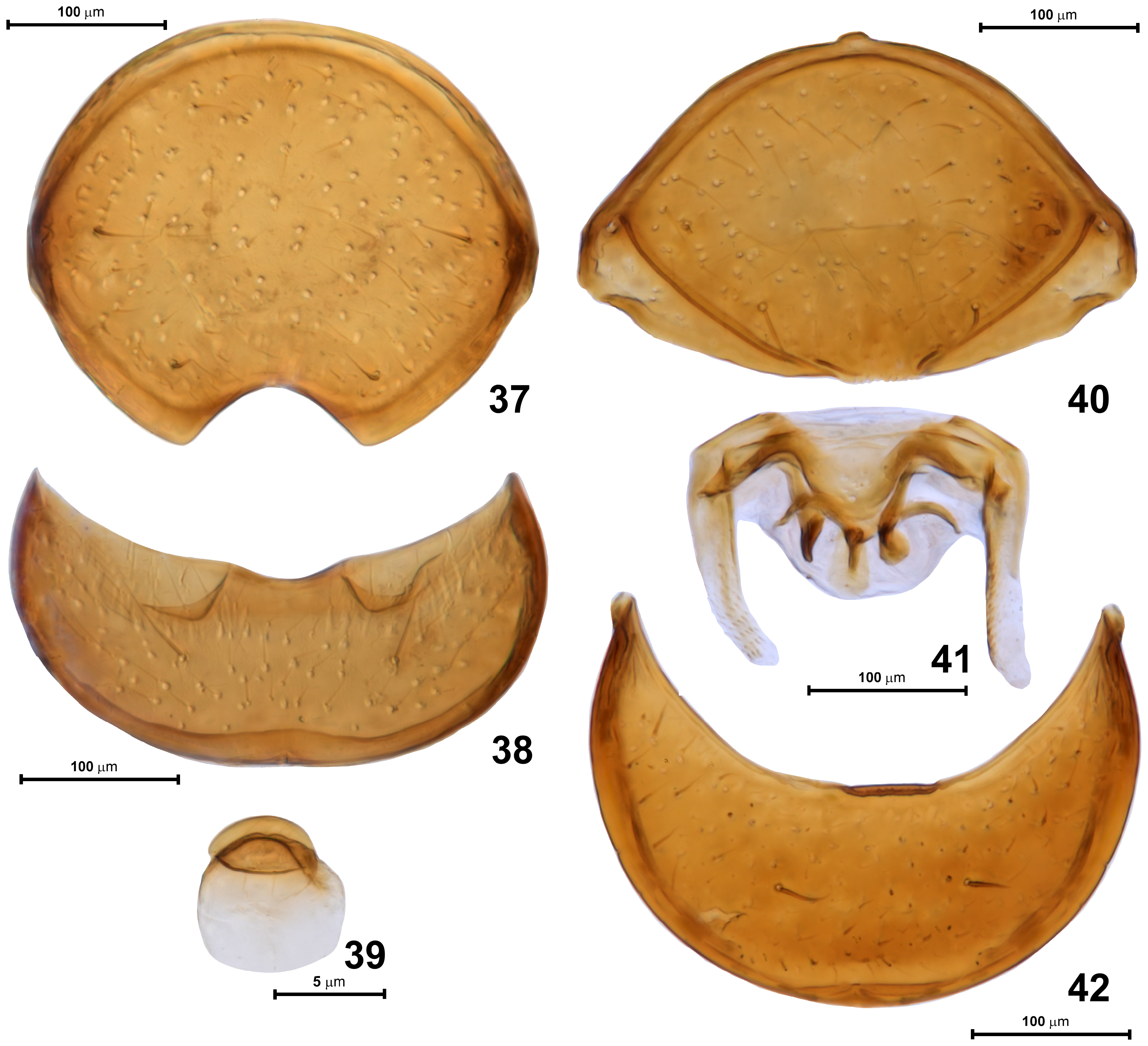
Abdomen ( Figs 27–39 View FIGURES 27–30 View FIGURES 31–36 View FIGURES 37–42 ) equal to or slightly longer than elytra, AbL 0.53–0.68 mm, AbW 0.60–0.75 mm. Tergite IV ( Figs 27, 28 View FIGURES 27–30 ) broadly subtrapezoidal, weakly narrowing posterad, convex in anterior half and strongly modified posteriorly; base with two pairs of foveae ( Fig. 28 View FIGURES 27–30 , indicated with arrowheads marked as “d”): one pair of shallow sublateral foveae directed mesad, with large setose openings ( Fig. 31 View FIGURES 31–36 ), and with median transverse impression filled with setae ( Fig. 32 View FIGURES 31–36 ) and flanked by pair of submedian foveae directed laterad, but lacking lateral discal carinae. Posterior half of tergite IV with large transverse impression subdivided into small anterior and much larger posterior cavities, the latter with large oval posteromedian tubercle ( Fig. 33 View FIGURES 31–36 ) sparsely covered with setae and surrounded laterally and anteriorly by asetose and smooth concave integument. In front of posteromedian tubercle, in small anterior part of impression, there are three pairs of slender cuticular processes: lateral process ( Fig. 34 View FIGURES 31–36 ; lp), submedian process ( Fig. 34 View FIGURES 31–36 ; smp), and median process ( Fig. 34 View FIGURES 31–36 ; mp). Lateral and median processes bear apical setae, each median process bears subapical and directed posteriorly penicillus composed of a few short setae ( Fig. 35 View FIGURES 31–36 ). Anterior margin of abdominal impression with strongly thickened cuticular walls (visible in cleared specimen: Fig. 28 View FIGURES 27–30 ). Median impression flanked by large and slightly impressed lateral setose patches ( Fig. 27 View FIGURES 27–30 ; lsp) composed of dense and short spatulate setae ( Fig. 36 View FIGURES 31–36 ), each seta with convex ventral/posterior and concave, grooved dorsal/ anterior surface, presumably forming glandular evaporation apparatus, with glandular opening situated anteriorly on setal papilla at base of each modified seta. Tergites V ( Fig. 30 View FIGURES 27–30 ) to VII short and each with pair of dorsal sublateral foveae ( Fig. 30 View FIGURES 27–30 ; indicated by arrowhead) directed posteromesad and slightly ventrad. Tergite VIII ( Fig. 37 View FIGURES 37–42 ) lentiform with deep arcuate posteromedian emargination and two pairs of long lateral setae.
Sternite IV ( Fig. 28 View FIGURES 27–30 , structures indicated with “v“, and Fig. 29 View FIGURES 27–30 ) with narrow keel-like intermetacoxal process ( Fig. 29 View FIGURES 27–30 ; imcp) and two pairs of foveae, all directed mesad: ventral sublateral foveae and ventral submedian foveae ( Fig. 28 View FIGURES 27–30 ; indicated with arrowheads marked with “v“). Sternites V–VII (not shown) narrow and lacking foveae; sternite VIII ( Fig. 38 View FIGURES 37–42 ) crescent-shaped with shallow posteromedian emargination and one pair of long setae; sternite IX ( Fig. 39 View FIGURES 37–42 ) minute, lentiform.
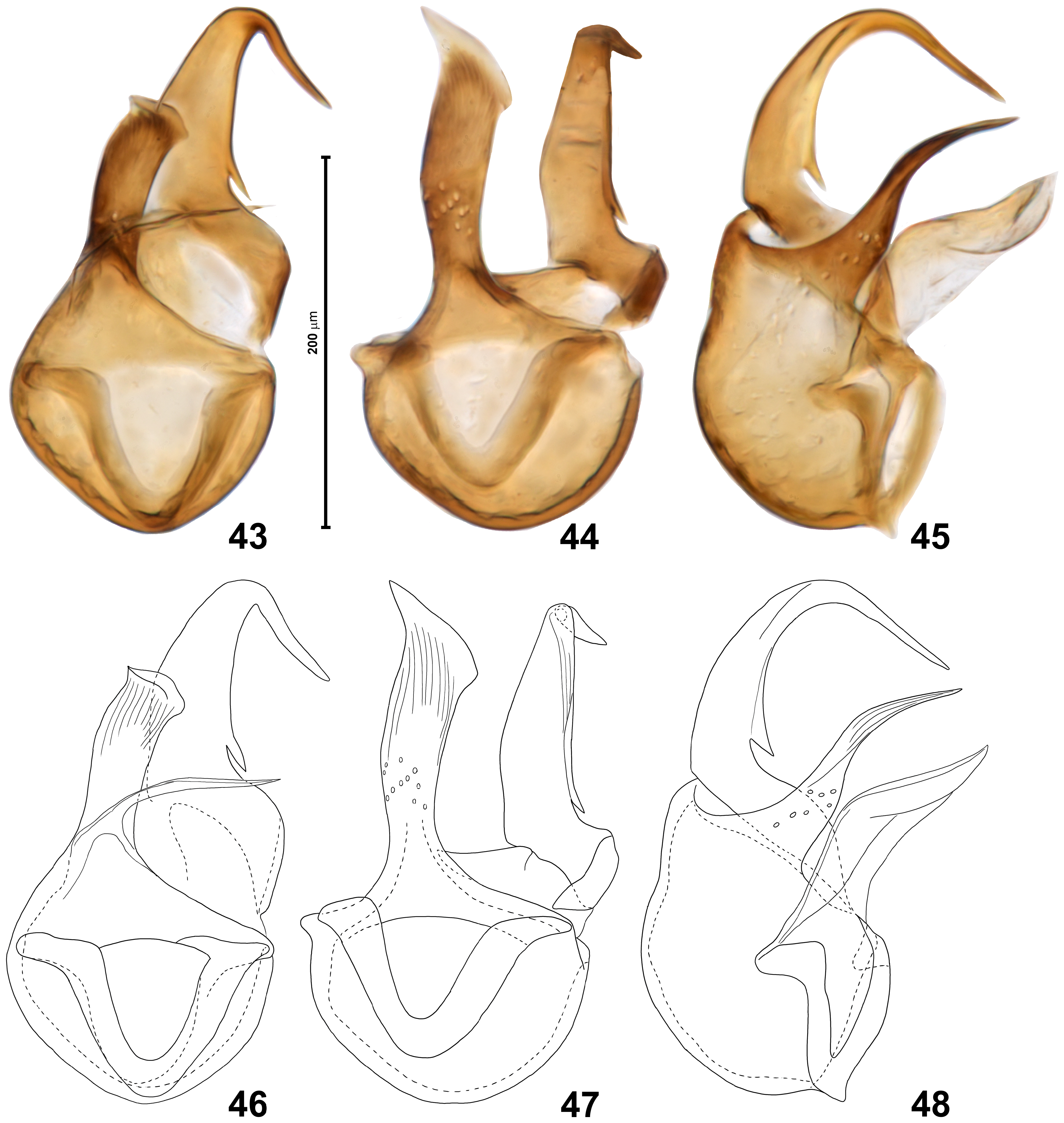
Aedeagus ( Figs 43–48 View FIGURES 43–48 ) with basal capsular region of median lobe stout, broader than long, ventral projection above ventral orifice slightly recurved, with subtriangular apex and strongly oblique apical margin; dorsal projection strongly bent ventrad at nearly right angle, with slander gradually tapering and pointed apex, proximal region of dorsal projection with small elongate subtriangular and pointed tooth directed proximally.
Female. Externally differs from male in unmodified abdomen with sternites IV–VIII exposed, unmodified porfemora, mesotibiae lacking apical mucro, and slightly shorter antennae in relation to body length. BL 2.08–2.13 mm; HL 0.43–0.45 mm, HW 0.44–0.45 mm, AnL 1.15 mm; PL 0.48 mm, PW 0.45–0.46 mm; EL 0.58 mm, EW 0.73–0.75 mm; AbdL 0.60–0.63 mm, AbdW 0.75 mm. Tergite VIII ( Fig. 40 View FIGURES 37–42 ) lentiform, posteriorly not emarginate; sternite VIII ( Fig. 42 View FIGURES 37–42 ) crescent-shaped with one pair of long setae and lacking posteromedian emargination; postabdominal sclerites ( Fig. 41 View FIGURES 37–42 ) complex, symmetrical (preparation in Fig. 41 View FIGURES 37–42 is slightly distorted).
Distribution. Okinawa Island, Ryȗkyȗ Archipelago, Japan.
Etymology. The new species is named in honor of Mr. Norihide Tokushige, who organized the field trip on which specimens of the first Arthromelodes known to inhabit Okinawa-jima were collected, and who himself discovered many interesting Pselaphinae and Scydmaeninae and significantly contributed to the knowledge of the Okinawan biodiversity.
Remarks. To date, eighteen species and two subspecies of Arthromelodes were known to occur in Japan ( Shibata et al. 2013). Fifteen of them inhabit the large ‘mainland’ islands (Honshȗ, Kyȗshȗ and Shikoku), one was discovered in Yakushima (~ 60 km S of Kyȗshȗ), and two on the northeastern Ryȗkyȗan Amami Islands. No species have been recorded from central or southwestern Ryȗkyȗs. When the identification key of Nomura (1991) is used, A. tokushigei may key out as A. crucifer Nomura or the pair A. sinuatipes Nomura and A. aizuanus Nomura , depending on interpretation of the punctures on head and pronotum (densely and coarsely punctate vs. sparsely punctate—it is difficult to decide which type of punctation better describes the condition in A. tokushigei ). However, none of the Japanese species revised by Nomura (1991) and later described by Arai (2002) has an even remotely similar aedeagus. Among the previously known Japanese species, only seven have the abdominal tergite IV modified: A. aizuanus Nomura , A. crucifer Nomura , A. dilatatus (Raffray) , A. optatus (Sharp) , A. saikaiensis Nomura , A. sinuatipes Nomura , and A. watanabei Arai. Arthromelodes aizuanus has the lateral setal patches on tergite IV strongly elongate, anteriorly extending far beyond the anterior margin of the posteromedian impression (in A. tokushigei not extending beyond the anterior margin of the impression); A. crucifer has the median impression on tergite IV elongate, with a “crossed carina“ ( Nomura 1991) and lacking oval posteromedian tubercle (in A. tokushigei there is no carina in the impression, which is posteriorly occupied by a large oval tubercle); in A. dilatatus the lateral setose patches on tergite IV are situated on broad and laterally protruding lobes (lobes lacking in A. tokushigei ); in A. optatus the impression on tergite IV bears a median longitudinal carina (lacking in A. tokushigei ); in A. saikaiensis the abdomen is strongly constricted at base (not constricted in A. tokushigei ); in A. sinuatipes the abdominal tergite IV has a peculiar shape, it is rapidly narrowed behind the lateral setal patches and the median impression is distant from the posterior tergal margin (tergite not rapidly narrowed and the impression posteriorly adjacent to the posterior tergal margin); and A. watanabei has strongly modified metatrochanters, each with a long curved ventral spine (lacking in A. tokushigei ).
Among Eastern Palaearctic species (including those inhabiting the transition zone between Palaearctic and Oriental realms), A. tokushigei somewhat resembles A. choui Yin, 2018 ( Taiwan) in the modification of the abdominal tergite IV and in the aedeagal structure. However, in A. choui the lateral setal patches on the abdomen extend anteriorly far beyond the posteromedian impression, and the latter is about as long as broad (in A. tokushigei the setal patches anteriorly do not extend beyond the anterior margin of the impression, which is clearly transverse), the pronotal punctures are inconspicous (in A. tokushigei distinct and dense), and the aedeagal projections are of different shapes.
Interestingly, among Batrisini of Okinawa-jima, females of A. tokushigei can be easily confused with females of Batriscenellus sakaii Nomura, 1991 . Both species were collected in the same spot, but B. sakaii is common also in other localities in the Yambaru area (Jałoszyński, personal obs.), while A. tokushigei has not been found by Japanese collectors who explored this island previously (no specimens have been seen by S. Nomura; email dated 26.04.2023). One of the reasons for overlooking A. tokushigei may be that this is a wingless species, so the commonly used flight intercept traps will not detect its presence. The females of A. tokushigei and B. sakaii have the same size and proportions of body parts; they clearly differ in distinct pronotal punctures in the former species vs. a smooth pronotal disc in B. sakaii , and in the structure of the postabdominal sclerites. As diagnoses of many genera of Batrisini seem extremely unclear (to the extent that they seem unusable) and this group will certainly require a profound reclassification, it is difficult to distinguish females of A. tokushigei and B. sakaii based solely on external generic features.
Specimens of A. tokushigei were collected by sifting leaf litter and rotten wood in a subtropical forest during a rather dry early spring, followed by extraction in a Winkler apparatus. Among many accompanying beetles found in the same sample of the substrate were Euaesthetinae : Stenaesthetus okinawaensis Puthz ; Paederinae : Nazeris okinawanus okinawanus Ito , Ochthephilum okinawaense Watanabe ; Pselaphinae : Morana deigo Arai , Morana kazuyoae Arai , Batriscenaulax kunigamensis Nomura , Batriscenellus sakaii Nomura , Tribasodites picticornis Nomura , Trisinus monostatos Nomura ; and Xantholininae : Diochus japonicus Cameron.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Pselaphinae |
|
Genus |