
Arctonyx albogularis, (BLYTH, 1853)
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2008.00416.x |
|
persistent identifier |
https://treatment.plazi.org/id/9D202A31-FFA0-FFFB-A41A-FCB5FDFA855F |
|
treatment provided by |
Felipe |
|
scientific name |
Arctonyx albogularis |
| status |
|
ARCTONYX ALBOGULARIS ( BLYTH, 1853) View in CoL
Taxonomic synonymy (unique names as originally proposed): Meles albogularis Blyth, 1853 .
Arctonyx taxoides Blyth, 1853 View in CoL .
Arctonyx isonyx Hodgson View in CoL , in Horsfield, 1856.
Arctonyx collaris taraiyensis Gray, 1863 View in CoL . ( nomen nudum).
Meles leucolaemus Milne-Edwards, 1867 View in CoL .
Meles (Arctonyx) obscurus Milne-Edwards, 1871 View in CoL .
Arctonyx leucolaemus orestes Thomas, 1911 View in CoL .
Arctonyx leucolaemus arestes Sowerby, 1914 . (nomen nudum; lapsus for orestes Thomas, 1911)
Arctonyx obscurus incultus Thomas, 1922 View in CoL .
Arctonyx leucolaemus milne-edwardsii Lönnberg, 1923 View in CoL .
Type material and type localities: The holotype of albogularis , from ‘Tibet’, presumably once in the collections of the Zoological Survey of India in Calcutta, was not listed among the type specimens still housed in that institution in 1977 ( Khajuria, Chaturvedi & Ghoshal, 1977). (We have examined one specimen from ‘Tibet’ – an adult skull, MNHN 1962-1641.) The holotype of taxoides is a subadult female, skin and skull, from ‘Assam’, deposited in the collections of the Zoological Survey of India in Calcutta ( Pocock, 1941: 429; Khajuria et al., 1977: addendum) The holotype of isonyx is apparently a young adult female skin at BMNH ( fide Pocock, 1941), with skull lost but redrawn by Pocock (1941) from Hodgson’s unpublished figures, from the ‘Terai of Nepal’ according to the original description and ‘Tibet’ according to Anderson (1879), but actually from the ‘ Sikkim Terai’ ( India) fide Pocock (1941). The nomen nudum ‘taraiyensis ’ was localized to the ‘ Sikkim Terai’ in the original description. The holotype of leucolaemus is MNHN 1866-89, a mounted specimen from the vicinity of Peking (= Beijing), China (G. Veron, in litt.). The holotype of obscurus is MNHN 1870-535, a mounted skin and accompanying skull, from Moupin, Sichuan, China (G. Veron, in litt.). The holotype of orestes is BMNH 11.6.1.6, a young adult female, skin and skull, from the ‘Tsing-ling Mountains, SW Shen-si’, Shaanxi Province, 12 000 feet, China. The holotype of incultus is BMNH 2.6.10.35, an old adult male, skin and skull, from ‘Chin-teh (Tsing-tö of Stieler), about 150 km W of Hang-chow’, Anhui Province, China. The holotype of milne-edwarsii is an adult female, deposited in the Swedish Museum of Natural History, from the Minshan Mountains of southern Gansu Province, China ( Lönnberg, 1923). Pei’s (1987) name Arctonyx minor , applied to a Pleistocene specimen from Guangxi Province, China, seems likely to be another synonym, but we have not seen the specimen on which the name is based or the original publication (published in Chinese; cf. Corbet & Hill, 1992: 199).
Common name: We suggest the common name ‘northern hog-badger’ for this species.
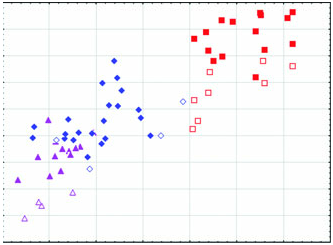
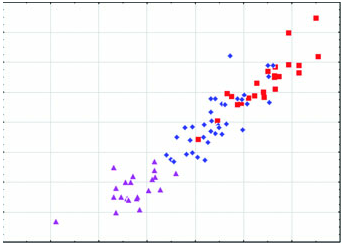
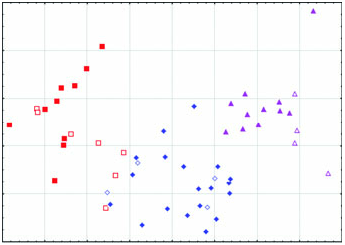
Diagnosis: Arctonyx albogularis is a markedly smaller badger than A collaris , considerably more gracile in all aspects of cranial conformation, and differs from that species in having smaller premolars and molars, on average ( Figs 2–6 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 : Tables 1–3). It differs from both A. collaris and A. hoevenii by attaining only a moderately developed sagittal crest in the oldest animals, in typically having a less pronounced diastema between P2 and P 3 in both the upper and the lower jaws, and in having a proportionally less expansive auditory meatus. Cranially, it differs further from A. hoevenii in having a larger skull on average, larger molars, a rostrum that is on average wider and proportionally less elongate, and a wider postdental palate ( Figs 2–6 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 ; Tables 1–3).
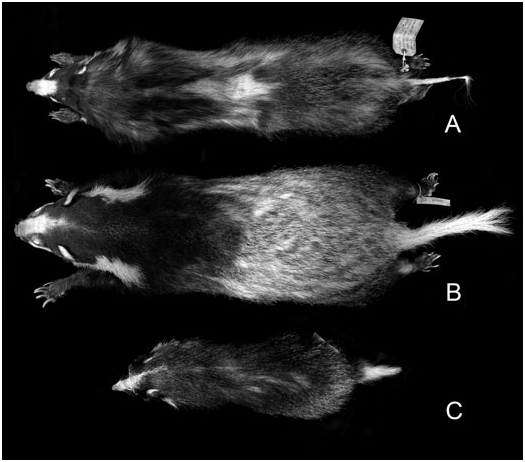
The pelage is softer and longer (the longer guard hairs in the winter pelage measuring ± 70 mm) than in other Arctonyx , with characteristically thick underfur in the winter months. The forequarters are blackish, and the mid-back, hindquarters and tail are white or heavily mixed with white. Although pelage coloration and marking varies somewhat geographically and individually, the back is more heavily overlaid with black guard hairs than in A. collaris , but less distinctly dark than in A. hoevenii ( Figs 9 View Figure 9 , 10 View Figure 10 ).
Distribution: The recorded historical distribution of Arctonyx albogularis extends from the southern foothills of the Himalayas (Sikkim; BMNH), Assam ( BMNH), Manipur (on the strength of a report by Rakamantha, 1994) and probably Bangladesh ( BMNH), across the Himalayas to Tibet ( Blyth, 1853; MNHN) and across the full expanse of southern and eastern China, from Gansu, Hebei, Shanxi and Liaoning Provinces in the north to Yunnan, Guangxi and Guangdong in the south ( Allen, 1938; Zhang, 1997). Records of Arctonyx from Nepal ( Shrestha, 1997) probably represent this species.
The distributional range of A. albogularis ( Fig. 11 View Figure 11 ) extensively abuts or overlaps that of A. collaris in the eastern Indian Subcontinent (e.g. in eastern India and perhaps Bangladesh), where both species are recorded ( Blyth, 1875; Blanford, 1888; Pocock, 1940, 1941; Rakamantha, 1994). Recently published reports of Arctonyx from the Indian states of Assam ( Choudhury, 1997a), Arunachal Pradesh ( Choudhury, 1997b), North [West] Bengal ( Choudhury, 1999) and Nagaland ( Choudhury, 2000), and from extreme northern Myanmar ( Rao et al., 2005) have not been identified to species, and could represent A. albogularis , A. collaris or both species in co-occurrence (cf. Rakamantha, 1994).
Arctonyx albogularis View in CoL is widely distributed and apparently common in much of China, its distribution extending over most of the eastern half of the country ( Zhang, 1997). Records of occurrence are from the provinces or municipalities of Liaoning, Hebei, Beijing, Henan, Shanxi, Shandong, Shaanxi, Ningxia, Gansu, Anhui, Hubei, Hunan, Jiangxi, Sichuan, Guizhou, Yunnan, Xizang ( Tibet), Zhejiang, Fujian, Jiangsu, Guangxi, Guangdong and Hong Kong ( Allen, 1929, 1938; Howell, 1929; Pocock, 1941; Zhang, 1997). Its modern or historical range is not known to extend into Mongolia or the Korean Peninsula, nor to the islands of Taiwan and Hainan. The southern limit of the species is not known. However, amongst three Arctonyx View in CoL skins without accompanying skulls, all taken at Lichiang, Yunnan, in early October 1916, one (AMNH 43095) is smaller and has the black back, longer and softer fur, and thick underfur typical of A. albogularis View in CoL , while two others (AMNH 43159, 43160) are larger, have paler backs, shorter, coarser fur and minimal underfur, and are thus probably referable to A. collaris View in CoL . Skulls would be needed to confirm the identity of these specimens definitively, but the characteristics of this series permit the suggestion that the boundary or area of overlap between the two species in southern China lies in Yunnan Province – a hypothesis in need of clarifying field investigations.
Arctonyx albogularis View in CoL seems to be a rather versatile species. Museum specimens document its occurrence across a remarkable elevational range, from sea level to at least 14 000 feet (i.e. 4300 m) in China. Although there are very few museum specimens available from the Himalayan region ( Pocock, 1940), Shrestha (1997) stated that in Nepal this species inhabits ‘forest and scrub in [the] elevational range [of] 1200–4000 m.’ Zheng et al. (1988) documented A. albogularis View in CoL in ‘forest-bush, farm land and wasteland, [and] mountain grassland’ in Shaanxi Province. Wang & Fuller (2003) found A. albogularis View in CoL to be relatively common in the vicinity of rural villages and surrounding agricultural landscapes.
Geographical variation: There is considerable variation in cranial size and shape within and between populations referred here to Arctonyx albogularis View in CoL . Much of this is individual (local) in nature, but strong geographical trends, particularly in size variation, are also apparent across the species’ range. We suggest that these are the principal reasons that so many geographical forms of A. albogularis View in CoL from China and India have been formally named by zoologists over the past 150 years (see above).
In his overview of the Chinese mammal fauna, Allen (1938) parsed geographical variation in A. albogularis (then called Arctonyx collaris ) taxonomically into two subspecies, ‘ A. c. collaris ’ of northeastern India and southern China (‘the hog-nosed badger of north-eastern India extends its range across southern China, apparently without important change of characters, not withstanding the various names that have been bestowed on it’; p. 403), and ‘ A. c. leucolaemus ’ of northern China, with records from Hebei, Gansu and the vicinity of Beijing. Later, Pocock (1940, 1941) elected to recognize the nominal Indian form taxoides as a subspecies distinct from Chinese races on the basis of its perceived smaller skull size. However, Pocock had very few mature specimens available for his comparisons (and indeed very few mature specimens of Indian A. albogularis remain available in museum collections today). We note that the skull of an adult Indian specimen of A. albogularis at RMNH (marked ‘Hindustan’, condylobasal length 142 mm) demonstrates that the range of size variation in Indian specimens (condylobasal length c. 110–142 mm) is broadly similar to that observed across all Chinese populations we have studied ( 116–149 mm). In addition to recognizing three subspecies of smaller, northern hog-badgers (i.e. A. c. collaris of the Himalayan region, A. c. albogularis of southern China and A. c. leucolaemus of northern China), Pocock (1941) drew from Allen’s (1938) tables of cranial measurements for Chinese hog-badgers to suggest that an additional, somewhat larger subspecies might be recognized from Fujian Province in south-eastern China.
Allen’s (1938) argument for recognition of a small northern Chinese subspecies of A. albogularis appears compelling on morphological grounds. Six adult specimens that we have examined from Beijing and Hebei (including older adults) are all very small, as is Lönnberg’s (1923) holotype of milne-edwardsii from Gansu (condylobasal length < 124 mm in all seven specimens). In northern animals the back of the head and forequarters tend to be white, whereas they are usually black in other populations of A. albogularis . Milne-Edward’s name leucolaemus was the first to be applied to this geographical sample (Milne- Edwards, 1867), and if leucolaemus is recognized as a subspecies, Lönnberg’s name milne-edwardsii can be regarded as a synonym ( Allen, 1938).
Populations of A. albogularis from central China are on average larger in cranial size than those from further north (we have examined specimens or series from Szechwan, Shaanxi, Zhejiang, Yunnan and Tibet). As noted above, Allen (1938) referred these to typical ‘ A. c. collaris ’, while Pocock (1941) recognized them under the name ‘ A. c. albogularis ’. As Allen (1938) and Pocock (1941) noted, specimens from Fujian Province are on average largest of all populations of A. albogularis so far studied.
In summary, a marked size gradient is apparent in Chinese A. albogularis , with animals on average being largest in south-eastern China, somewhat smaller in most of central China and relatively very small in northern China. Interestingly, this is the opposite of what would be predicted by Bergman’s Rule, a common scaling rule in mammals in which body size increases with increasing latitude (e.g. Meiri & Dayan, 2003; Meiri, Dayan & Simberloff, 2004). Pending further comparisons into the nature of this variation, we choose not to formally credit geographical variation in A. albogularis through the explicit recognition of subspecies, although both Allen’s (1938) discrimination of A. c. leucolaemus and A. c. collaris (i.e. here A. a. leucolaemus and A. a. albogularis ), and Pocock’s (1941) suggestion of additional subdivisions appear to have some merit. We advocate a combined study of molecular phylogeography, craniometric variation and pelage patterning across the wide range of this species in India and China to evaluate more incisively the significance of geographical variation within badger populations that we refer to A. albogularis . Amassing appropriate samples will require the assistance of colleagues especially in China, whom we encourage to deposit relevant vouchers in Chinese museum collections to complement those samples, documented in part here, that are currently available in other institutions.
Natural history: Arctonyx albogularis is a mediumsized badger (about the size of Meles meles ) that occurs in temperate forests and grasslands of eastern Asia (the Himalayas and China) at all elevations up to about 4300 m. It is an opportunistic omnivore (see below) and is apparently common in many areas. It lives in burrows, dug especially along rivers and streams and under boulders, and apparently is solitary except during the mating season in April and May ( Allen, 1938; Zheng et al., 1988). Young (litters apparently ranging in size from one to four) are born in February and March and weaned after about 4 months ( Zheng et al., 1988). Unlike other species of Arctonyx , A. albogularis hibernates throughout the winter, from November to February or May, at least in northern China ( Zheng et al., 1988). Zheng et al. (1988) reported two daily activity peaks for A. albogularis in Shaanxi, one at 03:00–05:50 h in the morning, and another at 19:00–2100 h in the evening. Northern hog-badgers are preyed upon by various large carnivores, including leopards ( Panthera pardus ), wolves ( Canis lupus ) and bears ( Ursus thibetanus ) ( Zheng et al., 1988).
We are aware of two studies on the diet of A. albogularis . Wang & Fuller (2003) collected 45 scat samples over the course of 1992–1994, in all months, near Taohang Village in Jiangxi Province, southeastern China. They found that A. albogularis was entirely animalivorous, feeding especially on small vertebrates (in 77% of samples, with rodents dominating), also taking a considerable proportion of gastropods (19% of samples), but no plant matter. The three other small to medium-sized sympatric carnivorans studied by Wang & Fuller (2003) ( Viverricula indica , Herpestes urva and Paguma larvata ) all fed on a greater proportion of plant material.
In considerable contrast, a detailed study of diet in A. albogularis at Long Xian in Shaanxi Province (based on dissections of stomachs from 57 animals) found that earthworms, roots, leaves, beetles, cicadas, lepidopteran larvae and acorns dominated in the diet, with some seasonal variability. Remains of small vertebrates (rodents, snakes, frogs and birds) were found in only 16% of stomachs. Earthworms were eaten by badgers with much greater frequency from late spring to autumn than in winter and early spring, and important earthworm taxa eaten included Allolobophora caliginosa, Pheretime hupeiensis, P. diffringens and Drawida japonica .
A few other additional, anecdotal observations on diet in A. albogularis are available. Long & Killingley (1983) noted that ‘two hog badgers in the Milwaukee Zoo ate “most anything,” both meat and vegetal.’ Gao & Sun (2005) described how Arctonyx diggings (for ‘insects and amphibians’) encourage seedling recruitment in Liaodong oaks ( Quercus wutaishanica ) in the Dongling Mountains of China. It seems clear from these various studies and accounts that A. albogularis is an opportunistic feeder, and that its diet no doubt varies with season, location and perhaps also with individual preference and with local differences in sympatric carnivoran assemblages.




| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Arctonyx albogularis
| Helgen, Kristofer M., Lim, Norman T-L. & Helgen, Lauren E. 2008 |
Arctonyx minor
| Pei 1987 |
Arctonyx leucolaemus milne-edwardsii Lönnberg, 1923
| Lonnberg 1923 |
Arctonyx obscurus incultus
| Thomas 1922 |
Arctonyx leucolaemus arestes
| Sowerby 1914 |
Arctonyx leucolaemus orestes
| Thomas 1911 |
Meles (Arctonyx) obscurus
| Milne-Edwards 1871 |
Meles leucolaemus
| Milne-Edwards 1867 |
Arctonyx collaris taraiyensis
| Gray 1863 |
Arctonyx taxoides
| Blyth 1853 |
Arctonyx
| F. CUVIER 1825 |
A. collaris
| F. Cuvier 1825 |
Arctonyx
| F. CUVIER 1825 |