
Hyoidea notaticeps Reuter, 1876
|
publication ID |
https://doi.org/10.5281/zenodo.5740129 |
|
publication LSID |
lsid:zoobank.org:pub:A5DF029E-037F-418D-BD77-BBC7C951592E |
|
persistent identifier |
https://treatment.plazi.org/id/9B7B87D3-FFF8-CF04-FE1F-E549FD3FFE6B |
|
treatment provided by |
Marcus |
|
scientific name |
Hyoidea notaticeps Reuter, 1876 |
| status |
|
Hyoidea notaticeps Reuter, 1876 View in CoL
( Figs 12–15 View Figs 10–17 , 18, 20, 27, 31, 34–36, 38, 47–48, 52, 62–63)
Hyoidea notaticeps Reuter, 1876: 34 View in CoL (original description)
Hyoidea notaticeps: KIRITSHENKO (1918: 152) View in CoL (record); KIRITSHENKO (1951: 181) (diagnosis); CARVALHO (1958: 76) (catalogue); HOBERLANDT (1963: 273) (redescription); KERZHNER (1964: 971) (diagnosis); PUTSHKOV & PUTSHKOV (1983: 17) (host); KAPLIN (1993: 166) (bionomics); QI et al. (1995: 59) (record); SCHUH (1995: 123) (catalogue); KERZHNER & JOSIFOV (1999: 249) (catalogue); LINNAVUORI & MODARRES (1999: 222) (record)
Type locality. Russian Federation, Volgograd prov., Sarepta [= Krasnoarmeysk district of Volgograd].
Type material examined. LECTOTYPE: ♀ (designated by HOBERLANDT (1963)), RUSSIAN FEDERATION: VOLGO- GRAD PROVINCE: Sarepta [= Krasnoarmeysk distr.of Volgograd], 48.5°N 44.48333°E, V.Jakovlev coll. (AMNH_PBI 00312702) ( ZISP). GoogleMaps
Additional material examined. HUNGARY: Budapest, 47.5004°N 19.02679°E, 122 m, 1700, unknown collector, 1 ♀ (AMNH_PBI 00340446) ( BMNH). R. Palota, 25 May 1885, G. Horváth, 1 J (AMNH_PBI 00340445) ( BMNH). KAZAKHSTAN:AKMOLA PROVINCE: Atbasar, 51.8°N 68.35°E, 30 Jul 1936, Rezvoy, 1♀ (AMNH_PBI 00312713) ( ZISP). ALMATY PROVINCE: Iliyskiy on Ili River, 43.52194°N 76.82972°E, 17 May 1937 – 18 May 1937, A. K. Lukyanovich,10JJ (AMNH_PBI 00312728–AMNH_PBI 00312737), 27♀♀ (AMNH_PBI 00312721–AMNH_PBI 00312724,AMNH_PBI 00312751–AMNH_PBI 00312766,AMNH_PBI 00312769–AMNH_PBI 00312771,AMNH_ PBI 00312773–AMNH_PBI 00312776) ( ZISP); 17 Jun 1937 – 18 Jun 1937,A.K. Lukyanovich, 2♀♀ (AMNH_PBI 00312767, AMNH_PBI 00312772) ( ZISP); 17 Sep 1937 – 18 Sep 1937, A. K. Lukyanovich, 1 ♀ (AMNH_PBI 00312768) ( ZISP), 1J (AMNH_PBI 00340288), 4♀♀ (AMNH_PBI 00340302–AMNH_PBI 00340305) ( ZMUH). Taldyqorghan [= Taldykurgan], 45°N 78.36666°E, 23 May 1937,A. K. Lukyanovich, 6♀♀ (AMNH_PBI 00308477, AMNH_PBI 00308478,AMNH_PBI 00312717–AMNH_PBI 00312720), 4JJ (AMNH_PBI 00308483,AMNH_PBI 00312655–AMNH_PBI 00312657) ( ZISP). KARAGANDA PROVINCE: Atasu [= Zhana-Arka], 48.68333°N 71.65°E, 18 Jul 1960,A.F.Emeljanov, 1♀ (AMNH_PBI 00312712) ( ZISP). SOUTH KAZAKHSTAN PROVINCE: Karasay,Dzhagan-ata, Kara-tau, 43.46666°N 69.51666°E, 27 May 1936 – 29 May 1936, A. K. Lukyanovich, 1 ♀ (AMNH_PBI 00311286) ( ZISP). WEST KAZAKHSTAN PROVINCE: Spartak on Bykovka River, 51.4°N 52.08333°E, 28 May 1949, K.G.Romadina, 1 ♀ (AMNH_PBI 00312714) ( ZISP). ZHAMBUL PROVINCE: Achi-Say,Teresakan river,Karatau Mts Ridge, 43.55417°N 68.88889°E, 01 Jun 1936, A. K. Lukyanovich, 2 ♀♀ (AMNH_PBI 00312710, AMNH_PBI 00312711) ( ZISP). RUSSIAN FEDERATION: ORENBURG PROVINCE: Nr Orenburg, 51.76666°N 55.1°E, 06 Jun 1924, A. I. Ivanov, 3 ♀♀ (AMNH_PBI 00312705–AMNH_PBI 00312707) ( ZISP). VOLGOGRAD PROVINCE: Sarepta [= Krasnoarmeysk], 48.5°N 44.48333°E, V. Jakovlev coll., 2 ♀♀ (AMNH_PBI 00312700,AMNH_PBI 00312701), 4 JJ (AMNH_PBI 00312651–AMNH_PBI 00312654) ( ZISP). SLOVAKIA: Čenkov, 47.95°N 18.53333°E, 07 Jun 1960, Exc. M. N. Pragae, Ephedra distachya (Ephedraceae) , 1J (AMNH_PBI 00340447), 1♀ (AMNH_PBI 00340448) ( BMNH); 05 Jun 1965, Štys, 1 J (AMNH_PBI 00340289), 5 ♀♀ (AMNH_PBI 00340295–AMNH_PBI 00340297,AMNH_PBI 00340293,AMNH_PBI 00340294) ( ZMUH); 08 Jun 1965, M.Kocourek,5JJ (AMNH_PBI 00337218–AMNH_PBI 00337222), 5 ♀♀ (AMNH_PBI 00337223–AMNH_PBI 00337227) ( NMPC). Štúrovo, 47.8°N 18.7333°E, 18 Jun 1991, H. Günther, 1 J (AMNH_PBI 00337217), 1 ♀ (AMNH_PBI 00337229) ( NHMM). TAJIKISTAN: Between Kvak and Kondara, Valley of Varzab River, 38.83°N 68.83°E, 17 May 1937 – 18 May 1937, A. K. Lukyanovich, 7 ♀♀ (AMNH_PBI 00312744–AMNH_PBI 00312750) ( ZISP); 08 Jun 1943,A. N. Kiritshenko, 2 larvae (AMNH_PBI 00312741,AMNH_PBI 00312742), 1♀ (AMNH_PBI 00312743) ( ZISP). TURKMENISTAN: Repetek, 38.58333°N 63.18333°E, 17 Apr 1905 – 18 Apr 1905, V.Oshanin coll., 1♀ (AMNH_PBI 00312703) ( ZISP); 29Apr 1972, Kaplin, Ephedra strobilacea (Ephedraceae) , 1J (AMNH_PBI 00312658) ( ZISP); 04 May 1972, Kaplin, Ephedra strobilacea (Ephedraceae) , 1 ♀ (AMNH_PBI 00312709) ( ZISP). UKRAINE: CRIMEA: Feodosiya distr., Karadag, 45.18°N 35.32°E, 07 May 1927, E. Kuznetsova and V. Kuznetsov, 1 ♀ (AMNH_PBI 00312708) ( ZISP). Kerch, 45.33333°N 36.45°E, 30 May 1915, A. N. Kiritshenko, 4 ♀♀ (AMNH_PBI 00312670–AMNH_PBI 00312672, AMNH_PBI 00312699) ( ZISP); 14 May 1917, A. N. Kiritshenko, 3 ♀♀ (AMNH_PBI 00312662–AMNH_PBI 00312664), 1 J (AMNH_PBI 00312633) ( ZISP); 17 May 1917, A. N. Kiritshenko, 2 JJ (AMNH_PBI 00308486, AMNH_PBI 00312648), 2 ♀♀ (AMNH_PBI 00312665, AMNH_PBI 00312666) ( ZISP); 19 May 1917, A. N. Kiritshenko, 1 ♀ (AMNH_PBI 00312667) ( ZISP); 21 May 1917, A. N. Kiritshenko, 1 J (AMNH_PBI 00312649) ( ZISP); 23 May 1918, A. N. Kiritshenko, 1 J (AMNH_PBI 00312650), 2 ♀♀ (AMNH_PBI 00312668, AMNH_PBI 00312669) ( ZISP). Katerlez [= Voykovo] nr Kerch’, 45.373°N 36.428°E, 09 May 1908, A. N. Kiritshenko, 2 ♀♀ (AMNH_PBI 00312660, AMNH_PBI 00312661) ( ZISP). 18 km NE of Voznesensk, Yuzhnyi Bug river, 47.694°N 31.489°E, 09 Jun 1921,A. N.Kiritshenko, 1J (AMNH_PBI 00312738) ( ZISP).Berdyansk N of Azov Sea Coast, 46.75°N 36.8°E, 15 May 1939, Topchiev, 1J (AMNH_PBI 00312726), 1♀ (AMNH_PBI 00312740) ( ZISP). Chongar, Sivash Lake, 46°N 34.5°E, 07 Jun 1940, A. K. Lukyanovich, 1 ♀ (AMNH_PBI 00308481), 2 JJ (AMNH_PBI 00308482, AMNH_PBI 00312725) ( ZISP). Dal’niye Makarty nr Berdyansk, 46.81666°N 36.63333°E, 22 May 1939, Nikolaev, 1 ♀ (AMNH_PBI 00312739) ( ZISP).Arnautka [= Kamyshany] nr Kherson, 46.61666°N 32.48333°E, 18 May 1939, Nikolaev, 1J (AMNH_PBI 00312727) ( ZISP). Kryzhanovka, NE of Odessa, 46.55°N 30.78333°E, 18 Jun 1924, E. Kiritshenko, 1♀ (AMNH_PBI 00312644) ( ZISP). Odessa, Khadzhib Liman, 46.46666°N 30.71666°E, 15 May 1921, A. N. Kiritshenko, 8 ♀♀ (AMNH_PBI 00308479, AMNH_PBI 00312634–AMNH_PBI 00312637, AMNH_PBI 00312640–AMNH_PBI 00312642), 6JJ (AMNH_PBI 00308484,AMNH_PBI 00312621–AMNH_PBI 00312625) ( ZISP); 28 May 1922, A. N. Kiritshenko, 3 ♀♀ (AMNH_PBI 00312638, AMNH_PBI 00312639, AMNH_PBI 00312643), 1 J (AMNH_PBI 00312626) ( ZISP). Odessa, Kuyalnitskiy Liman, 46.55°N 30.73333°E, 24 May 1923, A. N. Kiritshenko, 1 J (AMNH_PBI 00312627) ( ZISP). Odessa, Luzanovka, 46.46666°N 30.71666°E, 02 May 1920, A. N. Kiritshenko, 3 JJ (AMNH_PBI 00308485, AMNH_PBI 00312628, AMNH_PBI 00312629), 2 ♀♀ (AMNH_PBI 00312645, AMNH_PBI 00312646) ( ZISP); 04 May 1920, A. N. Kiritshenko, 2 ♀♀ (AMNH_PBI 00308480, AMNH_PBI 00312647), 3 JJ (AMNH_PBI 00312630–AMNH_PBI 00312632) ( ZISP).
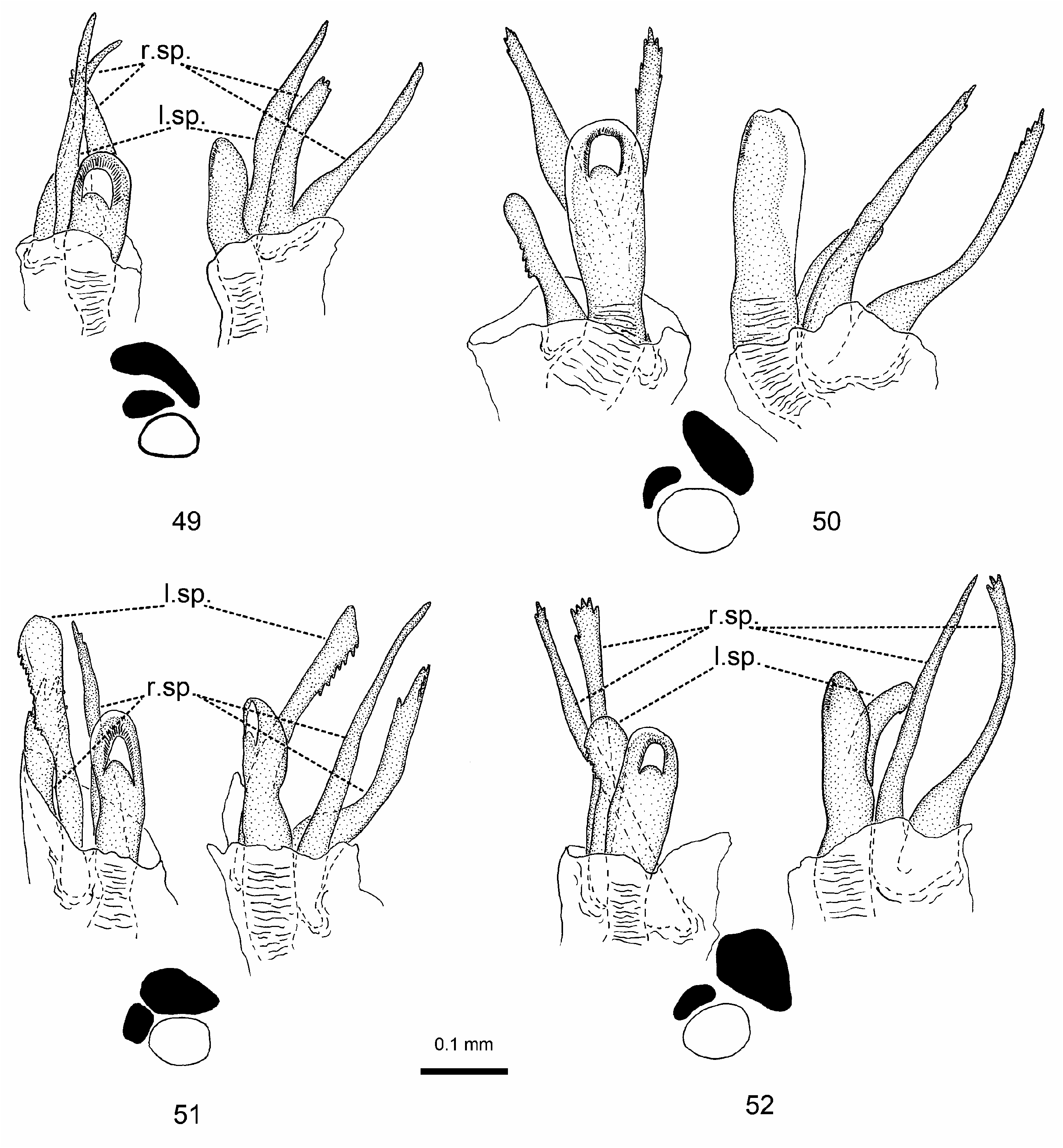
Redescription. Male: Small to middle-sized, 3.7–4.5 mm. COLOURATION ( Figs 12, 14 View Figs 10–17 ): Varies from dirty yellowish to brown. Head: Always pale brown; clypeus in pale specimens with black longitudinal spot at base and two black lateral stripes extending from base to middle of clypeus; in dark specimens clypeus entirely black; frons with black, confluent stripes radiating from midline, entirely merged in two large spots in dark specimens; vertex with two black, not confluent spots, in dark specimens with dark brown minute spots; antenna dark brown to black; labium brown to dark brown, apex of segment IV black. Thorax: Pronotal collar in pale specimens dirty yellowish with darkened central part, in dark specimens entirely brown; anterior part of pronotum dirty yellowish; calli covered with largely confluent black spots or entirely black; disc entirely yellowish in pale specimens, brown with more or less darkened base in dark specimens, sometimes with pale brown stripe along midline and pale basal edging; in some dark specimens disc at base with black minute spots; mesonotum pale brown to brown; scutellum dirty yellowish in pale specimens, dark brown to black with more or less expressed pale midline in dark specimens; thoracic pleurites dirty yellowish, sometimes with darkened central part; propleural suture black. Hemelytron: Clavus, corium and cuneus usually uniformly coloured, dirty yellow in pale specimens, brown in dark specimens, rarely corium in dark specimens with dark brown minute spots. Legs: In pale specimens, femora dirty yellow to pale brown, with a series of minute, partly confluent dark brown spots running along posterior margin at base and apically extending to anterior margin; in dark specimens fore femur additionally with large stripe along fore margin, middle femur with two large and partly confluent stripes along fore and hind margins, and hind femur almost uniformly brown to dark brown, with pale apex. Abdomen: Uniformly dirty yellowish in pale specimens, with more or less darkened tergites in dark specimens. SURFACE AND VESTITURE: Smooth, disc finely rugose in pale specimens and strongly rugose in dark specimens, with very fine punctures; scutellum typically with transverse wrinkles; vestiture composed of intermixed simple setae and silvery scales; simple setae uniformly pale in pale specimens, usually somewhat darker on hemelytron of dark specimens. STRUCTURE: Head: Vertex 2.4–2.8 × as wide as eye; antennal segment I 0.3–0.4 × as long as width of head; antennal segment II 1.1–1.5 × as long as width of head and 0.9–1.2 × as long as width of pronotum. Thorax: Pronotum 1.6–1.8 × as wide as long and 1.1–1.3 × as wide as head. GENITALIA: Genital capsule with large tooth on left side of genital opening ( Figs 31, 34–36 View Figs 30–36 ); sensory lobe of left paramere thick, curved and serrate ( Figs 47–48 View Figs ), caudal process apically dentate ( Fig. 48 View Figs ) or tooth-like, without denticles and with large claw-shaped additional process at base ( Fig. 47 View Figs ); right paramere as in Fig. 38; left spicule of aedeagus 0.5 × as long as right one, with serrate lateral margin; both branches of right spicule long and thin, with apical serration ( Fig. 52 View Figs 49–52 ).
Female: Small to middle-sized, 3.6–4.7 mm. COLOURATION ( Figs 13, 15 View Figs 10–17 ), SURFACE AND VESTITURE: As in male. STRUCTURE: Slightly larger than male, with larger interocular distance, vertex 3.0–3.6 × as wide as eye; antennal segment I 0.3 × as long as width of head; antennal segment II 0.9–1.1 × as long as width of head and 0.8–1.0 × as long as width of pronotum; pronotum 1.7–2.0 × as wide as long and 1.0–1.2 × as wide as head; abdomen larger and usually partly extending beyond apex of membrane; membrane reaching at least base of segment IX. GENITALIA: First gonapophysis as in Fig. 63; second gonapophysis as in Fig. 62.
Differential diagnosis. Distinguished by the following combination of characters: dark minute spots on dorsum absent; two black spots on vertex not confluent ( Figs 12–15 View Figs 10–17 ); simple setae on dorsum pale, rarely somewhat darkened on hemelytron; genital capsule with large tooth on left side of genital opening ( Figs 31, 34–36 View Figs 30–36 ); sensory lobe of left paramere thick, curved and serrate ( Figs 47–48 View Figs ), caudal process of variable shape ( Figs 47, 48 View Figs ); left spicule of aedeagus 0.5 × as long as right one, with serrate lateral margin; both branches of right spicule long and thin, with apical serration ( Fig. 52 View Figs 49–52 ). Most similar to H. lopezcoloni in the general colouration of dark forms and length of the antennal segment I, but differs in the shape of spicules of aedeagus and curved sensory lobe of left paramere.
Host associations and bionomics. According to HOBERLANDT (1963) and PUTSHKOV & PUT- SHKOV (1983), the host plants of this species are Ephedra distachya L., E. strobilacea Bunge , and E. intermedia Schrenk & C. A. Mey. The bionomics of H. notaticeps was carefully documented in the Repetek Nature Reserve, Turkmenistan ( KAPLIN 1993). The species was shown to be univoltine, larvae were collected from the end of March until the middle of May, adults – from April until May. According to HOBERLANDT (1963), in Central European conditions, adults occur from the middle of May until June.
Distribution. Widely distributed in the steppe zone and xeric habitats of Palaearctic (Fig. 18), in Slovakia and Hungary ( HOBERLANDT 1963), in southern Ukraine ( HOBERLANDT 1963), in Volgograd ( REUTER 1876, KIRITSHENKO 1951), Saratov ( KIRITSHENKO 1951) and Orenburg provinces ( KIRITSHENKO 1951) of Russia, in Georgia ( KIRITSHENKO 1918), Iran ( LINNAVUORI & MODARRES 1999), Kazakhstan ( HOBERLANDT 1963), Tadzhikistan ( KIRITSHENKO 1951), Turkmenistan ( PUTSHKOV & PUTSHKOV 1983), and in northern China ( Inner Mongolia: Tongliao and Hinggan) ( QI et al. 1995).
Discussion. The species shows considerable geographic variability in colouration and the structure of male genitalia. All specimens from the western part of its distributional range, particularly from Hungary and Slovakia are dark coloured, with largely confluent black stripes on the frons, and entirely black calli ( Figs 14–15 View Figs 10–17 ). All specimens from Kazakhstan are pale coloured, with non-confluent radiating stripes on the frons and calli with confluent dark spots but not entirely black ( Figs 12–13 View Figs 10–17 ). Both dark and pale morphs as well as specimens with intermediate colouration were found in the central part of the distributional area, i.e. in Ukraine and Volgograd province of Russia, with most specimens including the lectotype being pale. In terms of male genitalia, the western populations of H. notaticeps differ somewhat in having a blunt tubercle on the genital capsule ( Fig. 36 View Figs 30–36 ), similar to that of H. stehliki , and a single, apically serrate caudal process of the left paramere ( Fig. 48 View Figs ). In specimens from Kazakhstan the large tooth of the genital capsule is spine-like ( Fig. 34 View Figs 30–36 ) while the left paramere is equipped with an additional claw-shaped process at the base of the edentate caudal process ( Fig. 47 View Figs ). However, in both pale and dark specimens from southern Russia and Ukraine the shape of the large tooth of the genital capsule is somewhat intermediate ( Fig. 35 View Figs 30–36 ), while the left paramere bears a single caudal process. No differences in the aedeagus were revealed between western and eastern populations of H. notaticeps .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hyoidea notaticeps Reuter, 1876
| Knyshov, Alexander & Konstantinov, Fedor V. 2013 |
Hyoidea notaticeps : KIRITSHENKO (1918: 152)
| KERZHNER I. M. & JOSIFOV M. 1999: 249 |
| LINNAVUORI R. E. & MODARRES M. 1999: 222 |
| QI B. - Y. & LI W. - D. & LI SH 1995: 59 |
| SCHUH R. T. 1995: 123 |
| KAPLIN V. G. 1993: 166 |
| PUTSHKOV V. G. & PUTSHKOV P. V. 1983: 17 |
| KERZHNER I. M. 1964: 971 |
| HOBERLANDT L. 1963: 273 |
| CARVALHO J. C. M. 1958: 76 |
| KIRITSHENKO A. N. 1951: 181 |
| KIRITSHENKO A. N. 1918: ) |
Hyoidea notaticeps
| REUTER O. M. 1876: 34 |