
Agama, Daudin, 1802
|
publication ID |
https://doi.org/ 10.5281/zenodo.4665577 |
|
persistent identifier |
https://treatment.plazi.org/id/9A2EEA51-FFFE-FFC3-3FF2-14B81F9F5B3A |
|
treatment provided by |
Felipe |
|
scientific name |
Agama |
| status |
s.l. |
Agama s.l.
DESCRIPTION
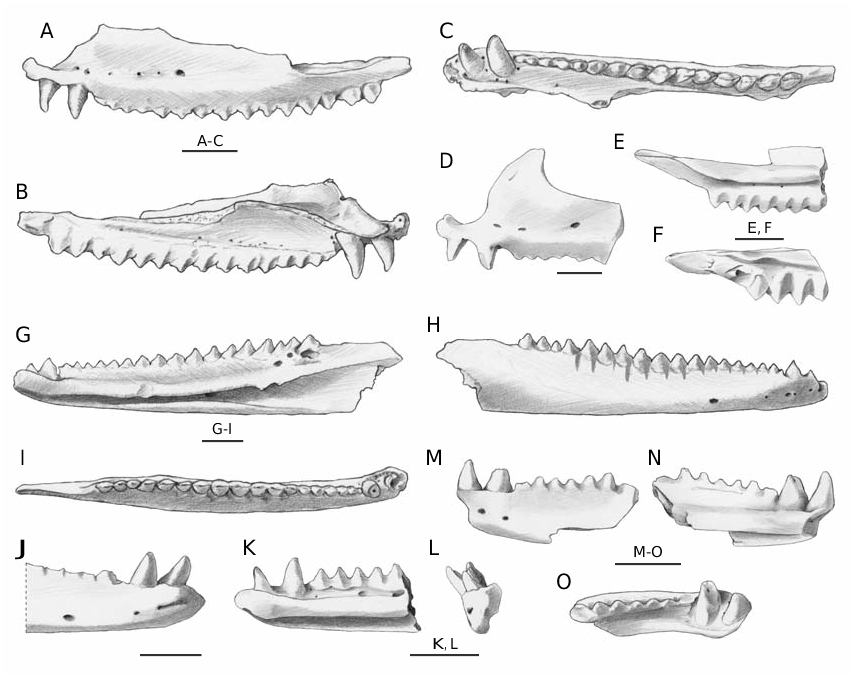
Maxilla
Due to the absence of a single complete maxilla, the description of the morphology of this element is possible thanks to the high number of fragments
or partly complete elements. The most complete
Fossil record and biogeography of European agamid lizards
maxilla, OR H VI 3/1 ( Fig. 2 View FIG A-C), is 16.3 mm long (the tip of the zygomatic process is broken off and therefore the original length was slightly higher). All the well-preserved maxillae are characterized by two anterior pleurodont teeth followed by at least 15 or 16 acrodont teeth.The orientation of the pleurodont teeth is approximately parallel to the external vertical wall while the acrodont tooth row is distinctly oriented medioventrally. The pleurodont teeth are located one anteriorly to and the other just under the beginning of the external vertical wall. They are well-spaced, monocuspidated, conical, pointed, slightly bent in posteromedial direction and, if well
preserved (like in the case of OR H VI 3/1), apically
provided with a mesiodistal keel.The acrodont teeth are not spaced (the posterior ones are even overlapped to the anterior part of the following tooth) and triangular in shape. The main cusp is in some cases flanked mesially and distally by a much smaller cusp.Teeth are provided with an evident mesiodistal keel which separates a nearly flat labial surface from a variably swollen lingual surface. The base of these teeth is slightly developed also on the medial surface of the bone and therefore should be better defined as pleuroacrodont ( Evans et al. 2002). The anterior acrodont teeth are small in size and variably worn (in one case they are nearly completely flattened by
usage) while the central and posterior ones are larger
Delfino M. et al.
(the size increases in backward direction), massive and partly overlapping: in several cases the posterior end of a tooth is labially overlapped to the anterior end of the following tooth (that is therefore lingually placed). The mesiodistal keel of the posterior teeth is therefore not parallel to the main axis of the tooth row but slightly inclined (anteromedially directed proximally and posterolaterally directed distally). Elongated wear facets are clearly visible on the lingual surface of all the acrodont teeth; these facets are always developed on both sides of the swollen lingual surface of each tooth and in some cases nearly reach each other toward the midline of the triangular tooth. There are no foramina along the acrodont tooth row but some foramina are present around the base of the pleurodont teeth. On the external surface, up to six foramina are aligned at the base of the prefrontal process; the largest is invariably the last one (usually corresponding to the sixth acrodont tooth). The prefrontal process is never entirely preserved. However, judging from the preserved portions, it can be stated that it is smooth, anteroposteriorly narrow but tall and that the anterior margin is usually rather steep ( Fig. 2D View FIG ) and subvertical at the base, at least in some cases. Above the line of foramina, a shallow anteroposteriorly depression is always present. The premaxillary process is distinctly developed not only in anterior but also in dorsal direction; if not damaged, the premaxillary process has a well-developed anterior surface for the suture with the premaxilla. On the dorsal surface of the external vertical wall, there is a distinct step at the end of the orbital margin (therefore corresponding to a step in the missing jugal). The zygomatic process is broken off in most of the cases; when present ( Fig. 2E, F View FIG ) it is long at least as the space occupied by the last four or five teeth. On the inner surface of the bone, the horizontal lamina is developed in mediodorsal direction reaching the maximum development approximately at the level of the sixth acrodont tooth. At least two foramina are developed in the groove produced by the dental shelf and the vertical wall.
Dentary
The dentaries are markedly slim and elongated, with a nearly straight ventral edge if seen in lateral view.
The best-preserved dentary OR H VI 3/5 ( Fig.2 View FIG G-I),
is 19.1 mm long but it is posteriorly broken off and anteriorly eroded. As for the maxillae, there are no complete dentaries and the above-mentioned specimen is the only one with an entirely preserved tooth row (length of the region with acrodont teeth is 12.9 mm). In this specimen, the two pleurodont teeth (partly broken off) are followed by 17 acrodont teeth. The shape of the pleurodont teeth (see also dentary fragments in Figure 2 View FIG J-O) does not differ from the one already described for the maxillary teeth; also the position and shape of the acrodont teeth are similar, but the lingual surface is less swollen than the labial one. The overlapping pattern is analogous, with posterior acrodont teeth a little labially overlapped to the following one. Another similarity concerns the wear facets but in the case of the dentary they develop considerably in ventral direction reaching the external bony surface of the dentary, forming evident long V-shaped grooves ( Fig. 2H View FIG ). The anterior acrodont teeth of the dentaries are approximately symmetric while the posterior ones are posteriorly inclined. In the few well-preserved dentaries, the anterior acrodont teeth are not completely worn as it happens in one fossil maxilla or in the dentaries of living agamids. On the medial surface, the symphyseal area is not clearly delimited but seems to have an oval shape. Few irregular foramina can be present at the base of the pleurodont teeth and in the longitudinal groove at the base of the acrodont tooth row. The Meckel’s groove opens along the entire length of the dentary: it is rather narrow in the anterior half of the dentary and slightly larger in the posterior half; under the last three or four teeth, the groove becomes narrower because of a blade-like ventrally directed expansion (partly broken in the specimen depicted in Figure 2G View FIG ) of the dorsal rim of the groove itself. The foramen for the inferior alveolar nerve (intramandibular foramen according to Bailon 1991) opens inside the groove, at the level of the 10th or 11th acrodont tooth. On the external surface, there are usually five aligned foramina distributed from the tip of the dentary to the level of the sixth or seventh acrodont tooth. The coronoid process, not completely preserved in any of the available dentaries, is dorsally thin
(blade-like), moderately long, gently sloping, but clearly posterodorsally oriented. There are no evident scars left by the coronoid. The angular process is invariably broken off.
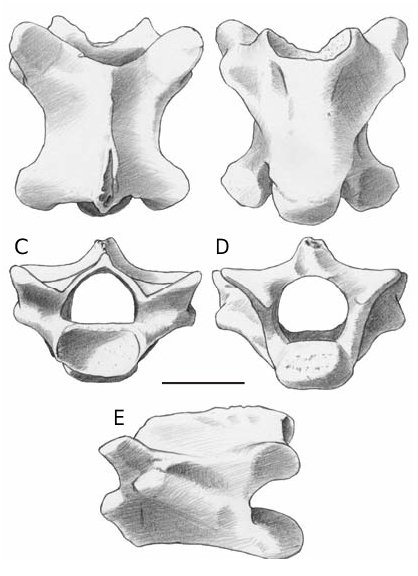
Vertebrae
Several vertebrae come from the cervical, dorsal and caudal region. All the vertebrae are procoelous.
Cervical and dorsal vertebrae ( Fig. 3 View FIG ). In anterior view, the cotyle is oval (dorsoventrally flattened). The neural canal is proportionally wide. The neural arch is triangular and devoid of zygosphene. The prezygapophyseal facets are dorsolaterally tilted. In dorsal view, the neural arch does not cover completely the centrum, of which the anteriorly convex edge is usually visible. The anterior edge of the neural arch is U- or V-shaped. Prezygapophyseal facets are oval-shaped and anterolaterally directed. The neural spine nearly reaches the anterior edge of the neural arch. Despite the presence of a moderate interzygapophyseal constriction, the neural arch has a rather wide appearance. The postzygapophyses surpass the posterior edge of the neural arch, which is modestly concave. In posterior view, the neural canal is wide. The neural arch is rather low and with straight lateral edges. The condyle has a shape that varies from oval to roundish. In ventral view, the dorsal rim of the cotyle overhangs the ventral one. The synapophyses are evident and laterally protruding. The centrum is triangular, anteroposteriorly compressed in the anterior trunk (cervical) vertebrae and more elongated in the posterior ones. The centrum ventral surface can be nearly flat or weakly convex, showing the presence of a modest, wide and smooth keel latero-anteriorly delimited by the hint of a groove. A couple of subcentral foramina can be present. In lateral view, the synapophyses are well defined, roundish or oval-shaped and placed at the level of the ventral rim of the cotyle or behind it. The ventral rim of the centrum is flat or moderately concave. Due to incompleteness of the neural spines, it is not possible to assess their dorsal development: the only exception shows a short neural spine of which the caudal edge does not surpass the posterior rim of the postzygapophyses. The few vertebrae with a hint of
hypapophysis located just behind the cotyle come
Fossil record and biogeography of European agamid lizards
A B
from the cervical sector. The maximum centrum length of a dorsal vertebra is of about 4 mm (but sensibly smaller on average).
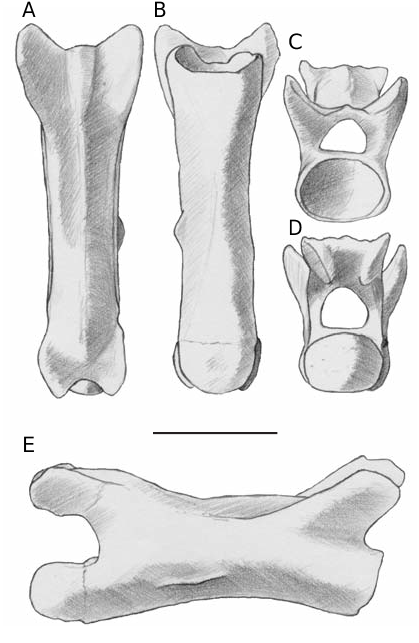
Caudal vertebrae ( Fig. 4 View FIG ). Caudal vertebrae are characterized by roundish cotyles and condyles, elongated centra devoid of autotomic plane and of fused chevron bones. The anterior caudal vertebrae preserve the proximal portion of robust and apparently long lateral processes (one per side), with a base dorsoventrally flattened, perpendicularly oriented to the main axis of the centrum and slightly pointing ventrally. The process-bearing vertebrae have a reduced neural spine represented by a ridge on most of the neural arch, but developed at the back of the neural arch as a robust spine pointing in dorsocaudal direction. The lateral processes and the
neural spine decrease in size in backward direction
Delfino M. et al.
along the tail and are not present in many of the fossil posteriormost vertebrae, where just a sagittal thin ridge is visible (as in the specimen depicted in Figure 4 View FIG ). The maximum centrum length of an anterior caudal vertebra is about 4.5 mm, the one of a posterior vertebra is about 5 mm.
REMARKS
The heterodont dentition characterized by pleurodont and acrodont teeth, unambiguously identifies the family Agamidae . Chameleons have a rather similar dentition but the anterior pleurodont teeth are missing and the acrodont teeth are usually more spaced, more labio-lingually symmetric (without an inflated surface) and located more apically than
those of agamids (for a discussion of the differences among these two groups see Moody & Rocek 1980; Bailon 1991; Blain 2005, and literature therein).The comparative osteology of agamids is incompletely known (the few exceptions are represented by Siebenrock 1895; El-Toubi 1947; Duda 1965; Ananjeva 1980, 1981, 1998; Moody 1980; Baig 1992) and the available information is hardly applicable to the fossil remains. Even if a confident allocation at genus level has to be considered as tentative at least, some considerations can be developed.
The referral of the remains from Monte Tuttavista to Phrynocephalus Kaup, 1825 and Trapelus Cuvier, 1817 , agamid genera presently inhabiting the periphery of Europe, can be excluded on the basis of remarkably different tooth numbers and/ or different morphology of the prefrontal process and external vertical wall (see Ananjeva 1980, 1981, 1998; Moody 1980).
Blain (2005) recently allocated at genus level ( Laudakia sp.) an agamid dentary from the late Pliocene of Vallirana ( Spain) mostly on the basis of the morphology of this element and acrodont teeth counts. The dentary from this locality possesses 14 teeth, a number which is congruent to that of the dentaries from Seynes and Medas islands (14 or 15 teeth) reported by Bailon (1991) and with that indicated by Baig (1992) for the Laudakia -group ( Stellio -group for the mentioned author), with the exception of L. tuberculata (Gray, 1827) that has 15-18 teeth. However, the teeth counts reported by Blain (2005: 243) for extant species do not match with those seen on the limited comparative material at our disposal. Blain reports the presence of 13 acrodont teeth in A. agama while the only specimen at our disposal (adult male from Kenya; M. Delfino pers. coll. no. 305) hosts 22 acrodont teeth. The morphological differences pointed out by Blain seem to be based on limited comparative material and not taking into account the ontogenetic variability. Cooper et al. (1970, and literature therein) report remarkable ontogenetic changes in terms of teeth counts (ontogenetic change from nine to 19 acrodont teeth in the dentaries of agamids) suggesting that this character can be applied only to large samples of fossil material and not to isolated findings. It is worth noting that the number
of acrodont teeth has not been included among the several cranial characters identified by Moody (1980) for the phylogenetic analysis of the agamids: the author explicitly writes that “characterization of agamid species and genera by number of acrodont teeth is difficult because of the radically differentsized anterior and posterior teeth and because of the enlargement of the anterior pleurodont teeth which erodes several of the anterior acrodont teeth” (Moody 1980: 62). The identification as cf. Laudakia sp. of the agamid fossils from another Spanish site, Casablanca-Almenara, reported by Blain (2005), is not based on any character that is exclusively present in the extant species of this genus; therefore such characters will be not further discussed here.
Moody (1980) considered as a valid character for his phylogenetic analysis of the agamids the position of the Meckel’s groove at the symphysis (if remains on the medial surface of the dentary – Stellio 57:0 – or if rotates to the ventral edge – Agama 57:1). Assuming that this character differentiates Agama from Laudakia , it could be used to identify the fossil remains from the Pliocene of Europe. According to his character matrix, all the analyzed species of Agama ( A. agama , A. atra Daudin, 1802 , A. hispida Kaup, 1827 ) have a Meckel’s groove which rotates ventrally, while all the analyzed species of Laudakia ( Stellio in his work; L. stellio , L. tuberculata ) have a groove which remains on the medial surface of the dentary (Moody 1980: appendix D). Surprisingly, on page 55 he states that “the groove rotates to the ventral edge as it rounds the symphysis in Agama [and] Stellio [= Laudakia ]”. The limited modern comparative material available for the present study does not show any significant difference among L. stellio (adult male from Corfu, M. Delfino pers. coll. no. 245), A. agama , A. bibroni Dumeril & Bibron, 1851 (adult specimen from Morocco, M. Delfino pers. coll. no. 275) and the few fossil dentaries from Monte Tuttavista that preserve the symphyseal region (a weak difference could concern the fact that in the fossils the symphysis is slightly more developed in medial direction). In all these cases, it seems that the groove is developed on the medial surface and never significantly rotates ventrally as
it approaches the symphysis.
Fossil record and biogeography of European agamid lizards
A further relevant character could be the orientation of the coronoid process of the dentary which is slightly posterodorsally oriented in the comparative modern specimen of L. stellio and in the few fossil remains from Sardinia, but slightly posteroventrally oriented in the comparative specimens of A. agama and A. bibroni . The orientation of this process has been also taken into consideration by Moody (1980; character 62; the process is called “retroarticular process”) but in the character matrix the status is reported to be the same for Agama and Laudakia (process projected posteriorly in a horizontal plane or curve dorsally; 62:0). Hence, the validity of this character for the identification of the members of these two genera should be confirmed on a wider sample of modern comparative skeletons (both in terms of number of species and of number of specimens for each species).
In conclusion, despite the fossil material from Sardinia here described resembles the only available comparative specimen of L. stellio in terms of maxillary and dentary acrodont teeth counts (the comparative specimen has 15 maxillary and 16 – or 17? – dentary acrodont teeth), shape of the coronoid process and position of the ventral lamina on the Meckel’s groove dorsal rim (that reduces the largeness of the groove posteriorly to the 14th acrodont tooth), a precise generic identification is here considered hazardous due to absence of any reliable osteological character allowing the distinction of Laudakia from Agama . The agamids from Monte Tuttavista are therefore formally referred to Agama s.l., which includes both these genera.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.