
Paranaitis
|
publication ID |
https://doi.org/10.1046/j.1096-3642.2003.00069.x |
|
persistent identifier |
https://treatment.plazi.org/id/793A8787-5C0E-6017-FF05-FF6EFC7FCFDD |
|
treatment provided by |
Carolina |
|
scientific name |
Paranaitis |
| status |
|
GENUS PARANAITIS View in CoL SOUTHERN, 1914
Paranaitis Southern, 1914: 66–67 View in CoL .
Anaitis Malmgren, 1865: 94 View in CoL . Junior homonym to Anaitis Duponchel, 1829 View in CoL (in Godart & Duponchel, 1829; Lepidoptera, Insecta).
Eulalia ( Euphylla) Knox, 1960: 113 .
Pareteone Hartmann-Schröder, 1975: 58–59 View in CoL .
Compsanaitis Hutching & Murray, 1984: 24 View in CoL .
Type species
Anaitis wahlbergi Malmgren, 1865: 94 View in CoL , pl. 14, fig. 31A–D, by monotypy.
Description
Prostomium anteriorly rounded, posterior part dorsally covered by first segment. Nuchal papilla may be indistinct, small and rounded, or larger and elongated, situated in posterior part of ligula (incision formed by segment 1). Nuchal organs retractile. Proboscis in some taxa subdivided into proximal and distal parts; lateral rows of papillae absent or present, and dorsal and ventral proboscis surfaces may be covered by chitinous or fleshy papillae and/or large tubercles. Segments 1 and 2 more or less fused, forming collar around prostomium. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered. Dorsal aciculae of segment 2 and 3 present or absent. Ventral aciculae from segment 2, chaetae from segment 2 or 3. Parapodia uniramous, dorsal cirri with rounded ends. Neuropodial lobes with supra-acicular lobe longer than subacicular. Rostrum of chaetal shafts symmetrical or asymmetrical, covered by many small teeth; single to few large main teeth present or absent. Ventral cirri oval, with rounded ends. Pygidium with single pair of oval or cylindrical pygidial cirri; single median pygidial papilla present.
Remarks
Euphylla was erected by Knox (1960) as a subgenus of Eulalia for his new species Eulalia ( Euphylla) benthicola and synonymised with Paranaitis by Pleijel (1991). The holotype and only known specimen has an unusually long and pointed nuchal papilla, in contrast to the rounded papilla in most Paranaitis species. As seen from, e.g. the presence of a collar and a well developed ligula, however, it clearly belongs within Paranaitis . A similar nuchal papilla is also found in P. caeca , from which it most notably differs in proboscis morphology (with dorsal papillae in P. caeca ; with rows of tubercles in P. benthicola ).
Hartmann-Schröder (1975) described a subfamily Pareteoninae and a genus Pareteone for her new species Pareteone abyssalis , known only by a single specimen. Pareteone was considered to be different from Paranaitis in having only three pairs of tentacular cirri on first two segments (1/1+1/N). Pleijel (1991) reexamined the holotype, and stated that the first pair of tentacular cirri are not absent but lost due to damage, and synonymized Pareteone with Paranaitis . Pareteoninae was consequently also synonymised with Phyllodocinae .
Hutching & Murray (1984) described the new genus and species Compsanaitis inflata , distinguished from Paranaitis by the lack of macropapillae on the proboscis. Paranaitis was characterized as having two lateral rows of papillae. Re-examination of Hutching & Murray’s specimens, however, shows that c. four macropapillae actually do occur on each side of the proximalmost part of the proboscis, and that rows of large tubercles are present on the distal part (their examination was probably based on specimens which did not have the proboscis fully everted). Accordingly, these, or any other observed characters, fail to justify a separation between Paranaitis and Compsanaitis , and we concur with Pleijel (1991) in regarding the latter name as a junior synonym.
PARANAITIS WAHLBERGI ( MALMGREN, 1865) View in CoL
( FIGS 1–3 View Figure 1 View Figure 2 View Figure 3 , 37 View Figure 37 )
Anaitis wahlbergi Malmgren, 1865: 94 View in CoL , pl. 14, fig. 31A–D; Grube, 1880: 214; Ditlevsen, 1909: 12; Fauvel, 1911: 26; Bergström, 1914: 155–156, fig. 51; Wesenberg-Lund, 1951: 25, 1953: 30.
Paranaitis wahlbergi View in CoL : Southern, 1914: 67, pl. 8, fig. 16; Hartman, 1959: 158–159, 1965: 62; Uschakov, 1972: 140–141, pl. 7, figs 8 and 9; Pleijel & Dales, 1991: 96, fig. 27A–C; Pleijel, 1993a: 30–32, figs 17,18,19, map 10.
Material examined
NORWAY: Holotype (SMNH-Type-233), Treurenberg Bay , Spitsbergen , Svalbard , 79∞56¢N, 16∞50¢E, 20–30 fathoms; 6 specimens (SMNH-22502), off Jan Mayen , 69∞01¢N, 8∞24¢W, 880 m; 5 specimens, 63∞10.0¢N, 04∞49.0¢E, 830 m, 21 March 1981, coll. T. Brattegard; 10 specimens, 63∞17.1¢N, 04∞24.8¢E, 1260 m, 22 March 1981, coll. T. Brattegard; 8 specimens, 65∞41.8¢N, 04∞22.9¢E, 1211 m, 7 June 1981, coll. T. Brattegard; 2 specimens, 62∞33.2¢N, 00∞58.9¢E, 800 m, 16 August 1981, coll. T. Brattegard; 7 specimens, 62∞33.6¢N, 00∞58.9¢E, 804 m, 21 January 1982, coll. T. Brattegard; 3 specimens, 63∞02.9¢N, 00∞48.5¢E, 1286 m, 15 August 1982, coll. T. Brattegard; 3 specimens, 63∞12.8¢N, 03∞07.3¢E, 1003 m, 23 August 1982, coll. T. Brattegard; 5 specimens, 63∞10.7¢N, 02∞45.9¢E, 1030 m, 26 November 1982, coll. T. Brattegard; 16 specimens, 62∞59.1¢N, 03∞13.1¢E, 804 m, 27 November 1982, coll. T. Brattegard; 1 specimen, 62∞07.4¢N, 04∞34.0¢E, 225 m, 27 November 1982, coll. T. Brattegard; 12 specimens, 65∞10.1¢N, 09∞29.6¢W, 784 m, 8 June 1983, coll. T. Brattegard; 5 specimens, 62∞35.6¢N, 01∞14.0¢E, 781 m, 17 June 1983, coll. T. Brattegard; 12 specimens, 62∞35.1¢N, 01∞47.6¢E, 656 m, 23 May 1984, coll. T. Brattegard; 2 specimens, 62∞17.6¢N, 01∞48.7¢E, 406 m, 21 November 1984, coll. T. Brattegard; 1 specimen, 62∞00.1¢N, 02∞01.3¢E, 374 m, 21 November 1984, coll. T. Brattegard; 23 specimens, 62∞31.5¢N, 01∞26.6¢E, 701 m, 8 January 1985, coll. T. Brattegard; 41 specimens, 62∞42.4¢N, 01∞11.2¢E, 897 m, 8 January 1985, coll. T. Brattegard; 12 specimens, 62∞54.7¢N, 00∞55.7¢E, 1112 m, 8 January 1985, coll. T. Brattegard; 1 specimen, 61∞14.4¢N, 02∞50.2¢E, 382 m, 23 March 1985, coll. T. Brattegard; 13 specimens, 60∞49.2¢N, 05∞22.3¢E, 81 m, 1 November 1985, coll. T. Brattegard; 2 specimens, 60∞49.2¢N, 05∞22.3¢E, 80 m, 1 November 1985, coll. T. Brattegard; 3 specimens, 63∞02.7¢N, 07∞01.7¢W, 1022 m, 13 June 1986, coll. T. Brattegard; 3 specimens, 69∞01.4¢N, 08∞24.6¢W, 876 m, 25 July 1986, coll. T. Brattegard; 23 specimens, 62∞50.6¢N, 01∞25.9¢E, 951 m, 15 August 1986, coll. T. Brattegard; 25 specimens, 62∞41.5¢N, 01∞45.4¢E, 750 m, 17 August 1986, coll. T. Brattegard; 2 specimens, 60∞49.2¢N, 05∞22.2¢E, 80 m, 17 October 1986, coll. T. Brattegard; 1 specimen, 60∞49.3¢N, 05∞21.6¢E, 85 m, 5 December 1986, coll. T. Brattegard. SWEDEN: 10 specimens, Säcken, Bohuslän, 125 m, 4 April 1988, coll. F.P. (mounted for SEM, not kept); 3 specimens (SMNH-23277), SW Y. Vattenholmen, Koster-area, Bohuslän, 160–180 m, 22 July 1988, coll. F.P.; 1 specimen (SMNH-22501), Singlefjord, Bohuslän, 80 m; 1 specimen (ZIHU-2014), Koster area. UNITED KINGDOM: 3 specimens ( NMW.Z 1985.023.0021), Scotland, Loch Creran, Argyll, 15– 22 m. GREENLAND: 1 specimen (ZUMC-POL-1500), Hurry Fjord, Fame Island, 16 August 1933, 22– 24 m.
Description
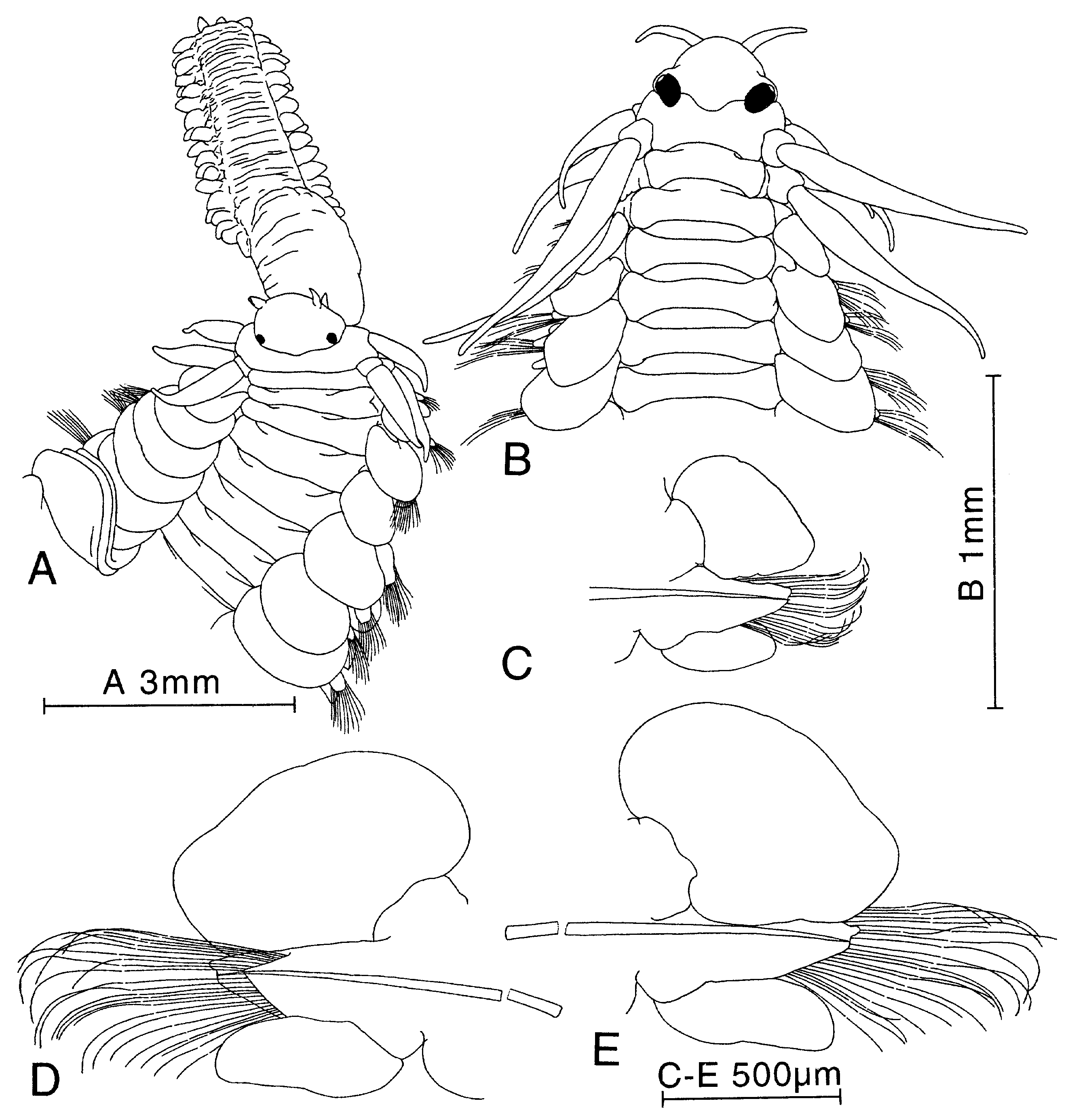
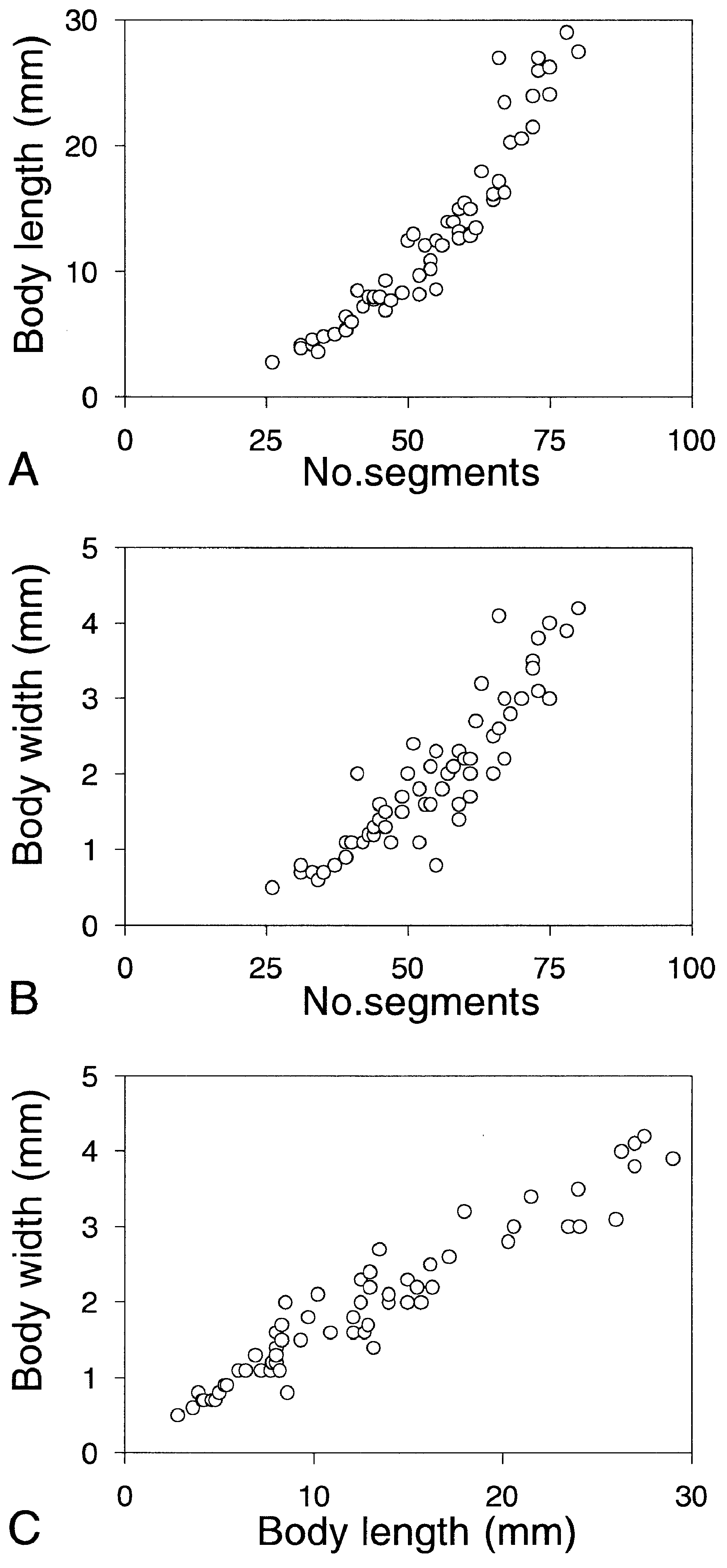
Holotype complete specimen, 73 mm long, 5.3 mm wide, for 110 segments. Up to 125 mm long, 15 mm wide, for more than 200 segments ( Uschakov, 1972); see Figure 3 View Figure 3 for measurements of other specimens. Live specimens whitish with clear red colour on dorsum and inner part of dorsal cirri from segments 5–6; eyes red. Preserved specimens whitish, larger specimens iridescent, eyes brownish black. Body dorso-ventrally flattened, almost of uniform width, with tapering posterior end. Prostomium anteriorly rounded, slightly wider than long, posteriorly covered by segment 1, with shallow ligula ( Figs 1A View Figure 1 , 2A,B View Figure 2 ). Paired antennae and palps short and narrow, anteriorly to laterally directed. Eyes rounded, with lenses, medium sized in holotype, partly covered by segment 1. Eyes proportionally larger in small specimens. Nuchal papilla indistinct to externally invisible ( Fig. 1A View Figure 1 ; see remarks). Nuchal organs retractile, knoblike, ventro-laterally situated. Proboscis with single lateral row of large fleshy papillae each side; each row with 3–5 merged lines of pointed papillae ( Figs 1B View Figure 1 , 2A View Figure 2 ). Terminal ring with large number of indistinct papillae, smaller on dorsal part than on ventral and lateral parts. Paired large papillae present laterally inside ring.
Segments 1 and 2 completely fused dorsally. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered distally. Cirri of segment 1 reaching segment 3–5. Dorsal cirri of segments 2 and 3 reaching c. segment 7–9. Ventral cirri of segment 2 reaching segment 4–6. Segment 2 without neuropodial lobes and chaetae. Segment 3 with neuropodial lobes with c. 3 chaetae; with small ventral cirri. Dorsal acicula of segments 2 and 3 absent.
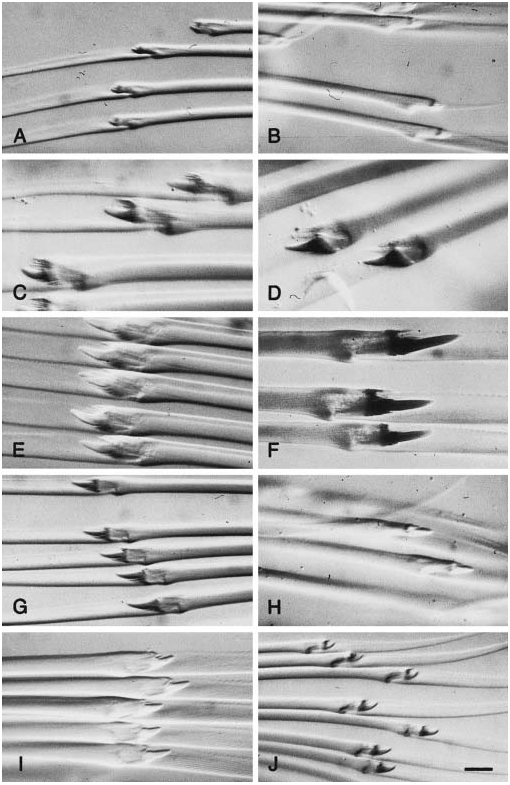
Dorsal cirri of median segments oval, asymmetrical, longer than wide ( Fig. 2C–E View Figure 2 ). Dorsal cirrophores short, indistinct. Neuropodial lobes long, with supraacicular lobes longer than subacicular lobes, with c. 75 chaetae in holotype, 20–30 chaetae in Swedish specimens ( Fig. 1C View Figure 1 ); more than 120 chaetae in large Arctic specimens. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, with 1–3 short main teeth on anterior side ( Figs 1D View Figure 1 , 37A View Figure 37 ). Ventral cirri elongated oval, with rounded ends, about as long as neuropodial lobes ( Fig. 2D View Figure 2 ). Pygidial cirri rounded, small, about as long as wide. Pygidial papilla present.
Habitat
Muddy bottoms, 10–1200 m. Remarks
Paranaitis wahlbergi differs from other Paranaitis in the character combination proboscis with paired lateral rows of rounded papillae, short and indistinct dorsal cirrophores, and rostrum of chaetal shaft possessing 1–3 short main teeth. It is similar to P. speciosa in proboscis and parapodia, but differs in pigmentation and chaetal morphology: P. speciosa has a greenish body and a single large main tooth on the rostrum of chaetal shaft.
Paranaitis wahlbergi View in CoL has been variously reported to have an indistinct nuchal papilla ( Uschakov, 1972) or no nuchal papilla ( Pleijel & Dales, 1991; Pleijel, 1993a). In their study of the central nervous system of phyllodocids, Orrhage & Eibye-Jacobsen (1998) recorded rudiments of a nuchal papilla, a rhomboid or triangular structure in cross-section situated just behind eyes. It was described as being innervated in the same way as the nuchal papilla of other phyllodocids. From our examination of a large number of specimens, a small indistinct projection was observed in most specimens at the position where a nuchal papilla is situated in other Paranaitis View in CoL , corroborating Orrhage and Eibye-Jacobsen’s anatomical observations.
Arctic and more southern specimens exhibit important size differences; whereas the former reach about 10 cm in body length, the latter do not exceed 2 cm. Furthermore, specimens from Sweden tend to have longer digitate cirri on the anterior-most segments, and less numerous chaetae. As both of these characters show a strong relationship to size, we nevertheless treat the Arctic and more southern populations as conspecific.

| T |
Tavera, Department of Geology and Geophysics |
| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Paranaitis
| Kato, Tetsuya & Pleijel, Fredrik 2003 |
Pareteone Hartmann-Schröder, 1975: 58–59
| Hartmann-Schroder G 1975: 59 |
Eulalia ( Euphylla )
| Knox GA 1960: 113 |
Paranaitis
| Southern R 1914: 66 |
Paranaitis wahlbergi
| Pleijel F 1993: 30 |
| Pleijel F & Dales RP 1991: 96 |
| Uschakov PV 1972: 140 |
| Hartman O 1965: 62 |
| Hartman O 1959: 158 |
| Southern R 1914: 67 |
Anaitis
| Malmgren AJ 1865: 94 |
Anaitis wahlbergi
| Malmgren AJ 1865: 94 |
Anaitis wahlbergi
| Wesenberg-Lund E 1951: 25 |
| Bergstrom E 1914: 155 |
| Fauvel P 1911: 26 |
| Ditlevsen H 1909: 12 |
| Grube AE 1880: 214 |
| Malmgren AJ 1865: 94 |