
Nyctimene wrightae, Irwin, 2017
|
publication ID |
https://doi.org/10.3853/j.2201-4349.69.2017.1654 |
|
persistent identifier |
https://treatment.plazi.org/id/706DD361-D448-FFF6-FED5-AE819B77CC0A |
|
treatment provided by |
Felipe |
|
scientific name |
Nyctimene wrightae |
| status |
sp. nov. |
Nyctimene wrightae View in CoL sp. nov.
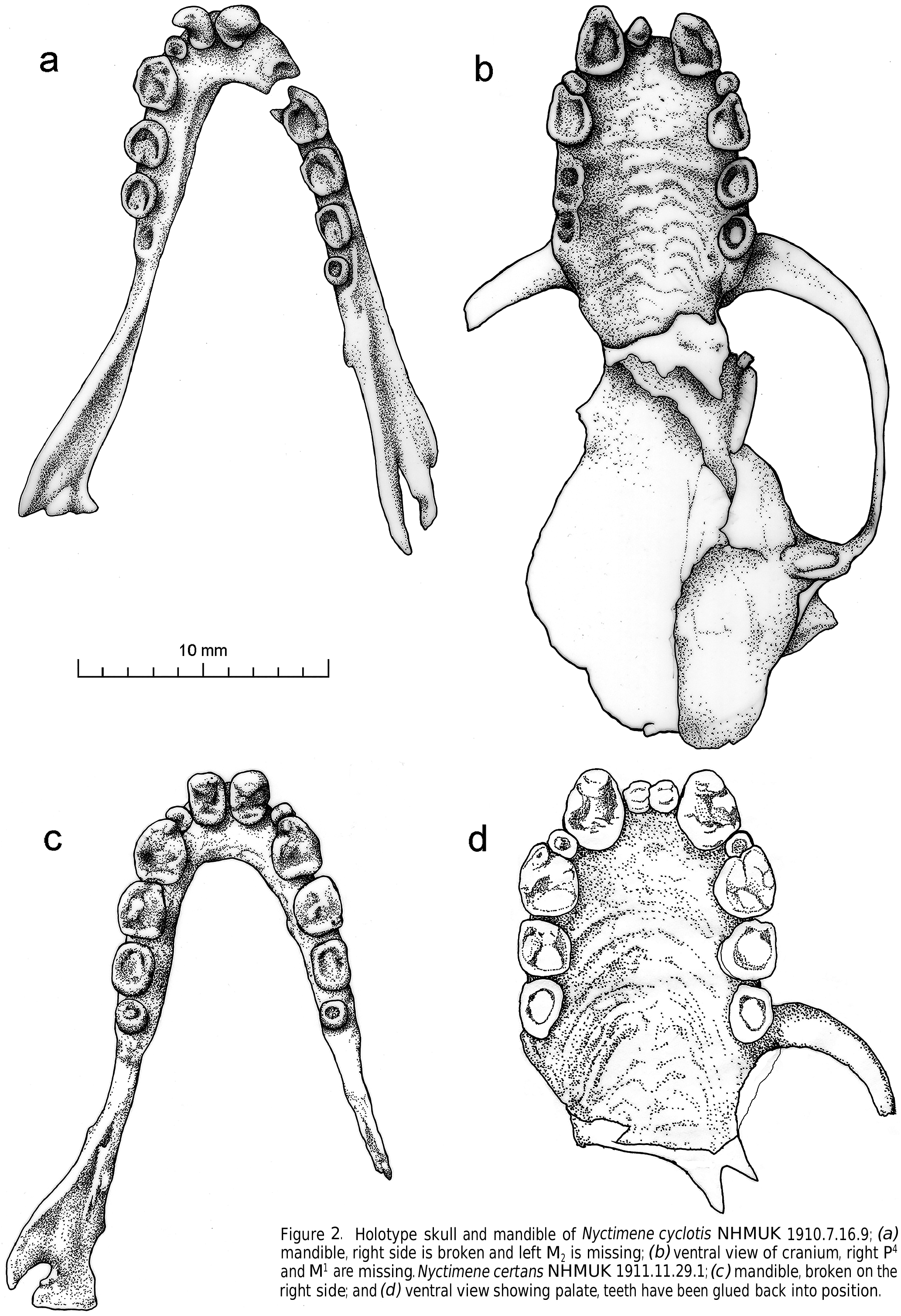
Figs 3 View Figure 3 , 4 View Figure 4 , 6–12 View Figure 6 View Figure 7 View Figure 8 , Tables 2–6
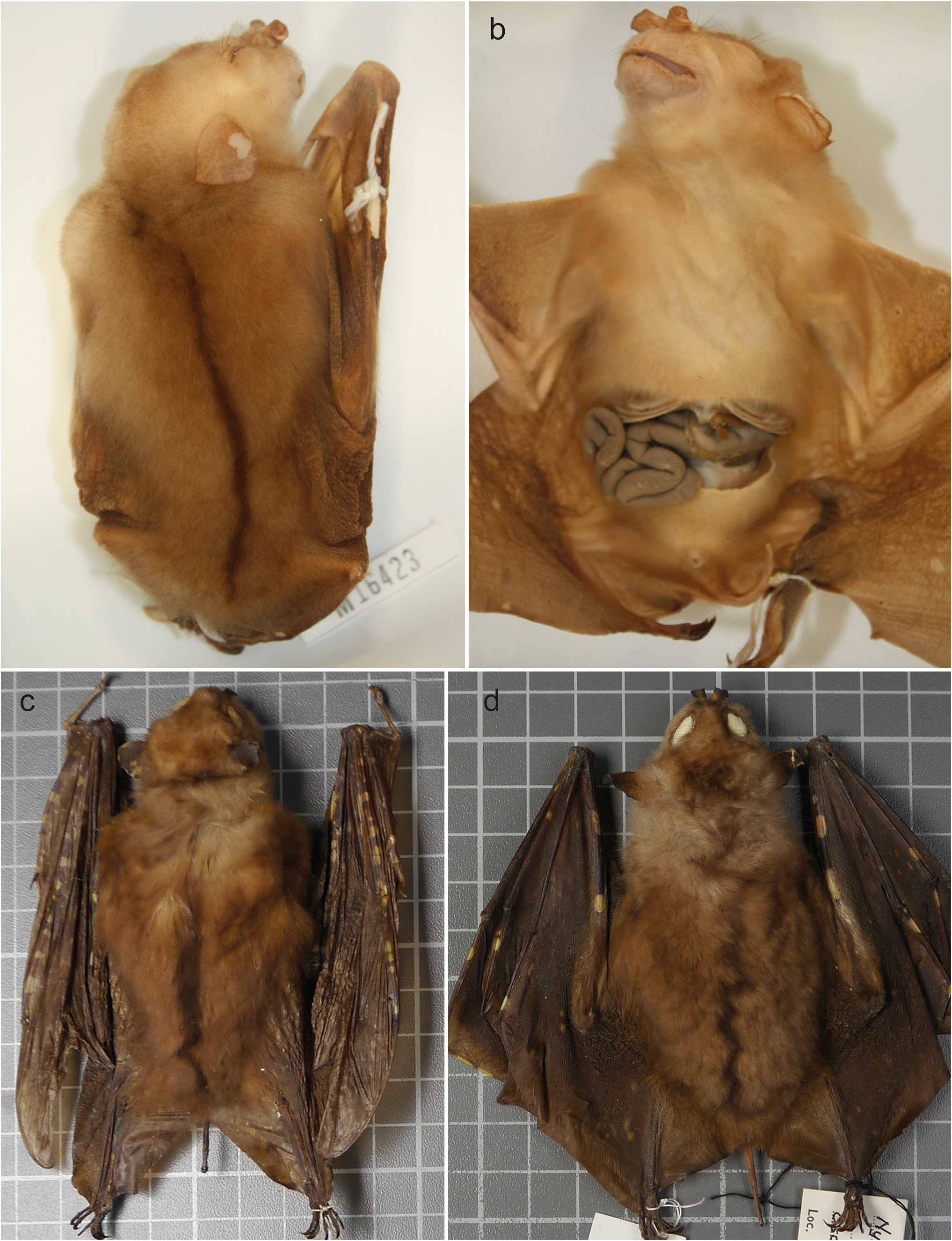
Holotype. AM M.16423, ♀, adult, whole body in spirit, skull extracted, collected 7 Nov. 1985 by Dr Ken Aplin . Paratypes: A total of 29: AM M.16230 ( Field number FA 332 ), ♂ spirit and skull , AM M.16231 ( Field number FA 357 ), ♀ adult, spirit and skull, both collected on 08- April-1986 , AM M.16870 (field number FA 394 ), 1♂ adult, spirit body, collected 09-Apr-1986, all three collected by T. Flannery and T. Ennis from Munbil , Star Mountains, West Sepik Prov., PNG . AM M.16426 (field number X19), 1♂ adult, spirit and skull collected 22-Nov-1985 , AM M.16428 (field number X97), 1♀ adult, body in spirit, collected 24- Nov-1985 , AM M.16432 (field number Y20), 1♂ adult, body in spirit, collected 25-Nov-1985, all three collected by K. Aplin from Waro , Southern Highland Prov., PNG . AM M.16421 (field number H29), 1♀ sub-adult, collected 17-Oct-1985 , AM M.16422 (field number H49), 1♀ adult, collected 19-Oct-1985, both bodies in spirit and skulls extracted , AM M.16440 (field number G97), 1♀ adult, body in spirit, collected 15-Oct-1985 , AM M.16441 (field number L24), 1♀ adult, spirit body, collected 14-Oct-1985, all four collected by K. Aplin, Magidobo , Southern Highland Prov., PNG . AM M.16443 (field number F04), 1♂ adult, spirit body and skull, and AM M.16444 (field number K10), 1♀ adult, spirit body, collected respectively 7 and 11-Oct-1985, by K. Aplin in Bobole village, Mt Sisa, Southern Highland Prov., PNG . AM M.17887 (field number FB2), 1♂ adult, skin and skull, collected 30-Mar-1986 by T. Flannery in regrowth, outskirts of Tabubil , Western Province, PNG . AM M.21771 (field number FJ139 ), 1♀ sub-adult, body in spirit, collected 08-Mar-1990 by T. Flannery, Mt Somoro Summit , West Sepik Prov., PNG . AM M.32382 (field number B32), 1♂ adult, skin and skull, collected 12-Jun-1994 by Boeadi, Timika area , roadside forest at new town complex, Papua Prov., Indonesia . BPBM 24539 About BPBM , 1♂ adult, skin and skull collected 5-April-1965 and BPBM 24570 About BPBM , 1♂ adult, skin and skull, collected 10-Apr-1965 by P. J. Shanahan, Sinaeada , Milne Bay Prov., PNG . BPBM 28396 About BPBM , 1♀ adult, skin and skull, collected 10-Jan-1967 by P. H. Colman, Mt Missim ( S Slopes), Morobe Prov., PNG . BPBM 51380 About BPBM , 1♀ adult, skin and skull, collected 05-Aug-1967 by P. H. Colman, 10 km W Bulolo , Morobe Prov., PNG . BPBM 99227 About BPBM , 1♀ adult, skin and skull, collected 02-Oct-1970 by F. J. Radovsky, Bupu river 12 mi, NE Lae, Morobe Prov., PNG . BPBM 103790 About BPBM , 1♀, spirit, collected 16-Feb-1974 by A. B. Mirza from 5 km N of Wanuma , Madang Prov., PNG . BPBM 109893 About BPBM , 1♂ adult, skin and skull, collected 24-Sep-1986 by A. E. Engilis Jr, from 8.5 km N, 13 km E Utai, Agpo Creek ( Camp I), West Sepik Prov., PNG . NHMUK 1969.1417 About NHMUK , 1♂ adult, skin and skull, collected 14-Jul-1969 by J. I. Menzies from Olsobip , Upper Fly, Western Prov., PNG . PMAG 25845, 1♀ adult, spirit specimen collected from Lababia, Morobe Prov., PNG . PMAG 25847, 1♀, PMAG 25849, 1♂, and PMAG 25851, 1♂, all adult spirit specimens, collected from Aelalar river , Kawiali, Morobe Prov., PNG . WAM47463, 1♀ adult, and WAM47464, 1♂ adult, bodies in spirit and skulls extracted, collected 01-Jan-1997 by D. J. Kitchener, Timika , Papua Prov., Indonesia .
Specimens examined. A total of 119, vouchered bodies 32, vouchered and field data from 87 released individuals as listed below: PMAG un-catalogued ♂♂, adults spirit specimens (NM33 and NM 35 in my database), collected from Tamerce, Mt Loves , Hirtano Highway, Central Prov., PNG. DW2085 1♀ adult sampled on 05-Nov-1996 and DW2089 1♀ adult, and DW2090 1♂ adult, sampled on 16-Nov-1996, field data & tissues provided by Debra Wright, Ivimka, Gulf Prov., PNG. NRI 117, 312, 411, 622, 689, 712, 721, 729, 736, 781, OM11, OM 88 adult ♀ ; NRI 007 ♂, measured 138b, 683, 706, and released 715, 752, all adult ♂♂, NRI 722 sub-adult ♂, from Sempi, Madang Prov., PNG by Nancy Irwin 1996–2000. NRI 261, 287, 299, 311, 889, 892–893, 900, 903 all sub-adult ♀♀ ; NRI 307, 814, 855, 882, sub-adult ♂♂; NRI 241, 255, 263, 279, 291–292, 306, 763, 766–767, 769–70, 773–775, 780, 782–786, 795, 798, 801, 803, 805, 808, 815, 831, 838, 850, 852, 858–859, 879, 886, 897, 902, 910 adult ♂♂; NRI 252, 262, 288–289, 300, 776, 809–810, 823, 865, 867, 877, 880, 895, 914, adult ♀♀ measured and released from O-Pio, Chimbu Prov., PNG by Nancy Irwin 1999–2000 .
Type locality. Namosado ( 06°15'S 142°47'E, 500 m asl), Southern Highlands Province, PNG GoogleMaps .
Distribution. Widespread from lowland to montane forest of New Guinea.
Diagnosis. Nyctimene wrightae sp. nov. differs from all other species of Nyctimeninae in having the shortest mean rostrum length (R-L) relative to skull length (GSL) (<0.8); and differs from all Nyctimeninae other than N. certans and N. cyclotis , in having a proportionately broader dental arcade and palate as reflected by a P 3 -P 3 that is distinctly greater than P 1 -M 1, compared to a P 3 -P 3 being equal or shorter than P 1 -M 1 in other species of the genus. It differs from all species of the genus, other than N. certans , in having the greatest Zygo-B relative to skull length (> 79% compared to <74%). It differs further from all other species of the genus, other than N. certans and N. cyclotis , in its reduced width across M 1 -M 1 compared to P 4 -P 4.
Nyctimene wrightae sp. nov. is further distinguished from N. certans and N. cyclotis in having short (< 10 mm), brown, dorsal fur versus grey-brown fur, as well as a clearly defined brown dorsal stripe, compared to the ill-defined and often indistinct dorsal stripe of N. certans and N. cyclotis ; the basal portion of the hair shaft that is dark brown is usually less than 10%, with 5–10% of the distal portion of the hair is dark similar to that of N. a. papuanus , in sharp contrast to much higher proportion of dark fur of N. certans and N. cyclotis ; N. wrightae sp. nov. differs further in having ears that are longer than broad and which taper towards the apex typical of most Nyctimene species , in contrast to the short, broad ears of N. certans and N. cyclotis
Nyctimene wrightae sp. nov. is readily confused with the sympatric N. a. papuanus in external appearance but the latter species is of smaller average size. Nyctimene wrightae sp. nov. is readily distinguished by its smaller size, e.g., FA 52–63 mm, compared to FA> 67 mm for N. aello , N. major and N. rabori .
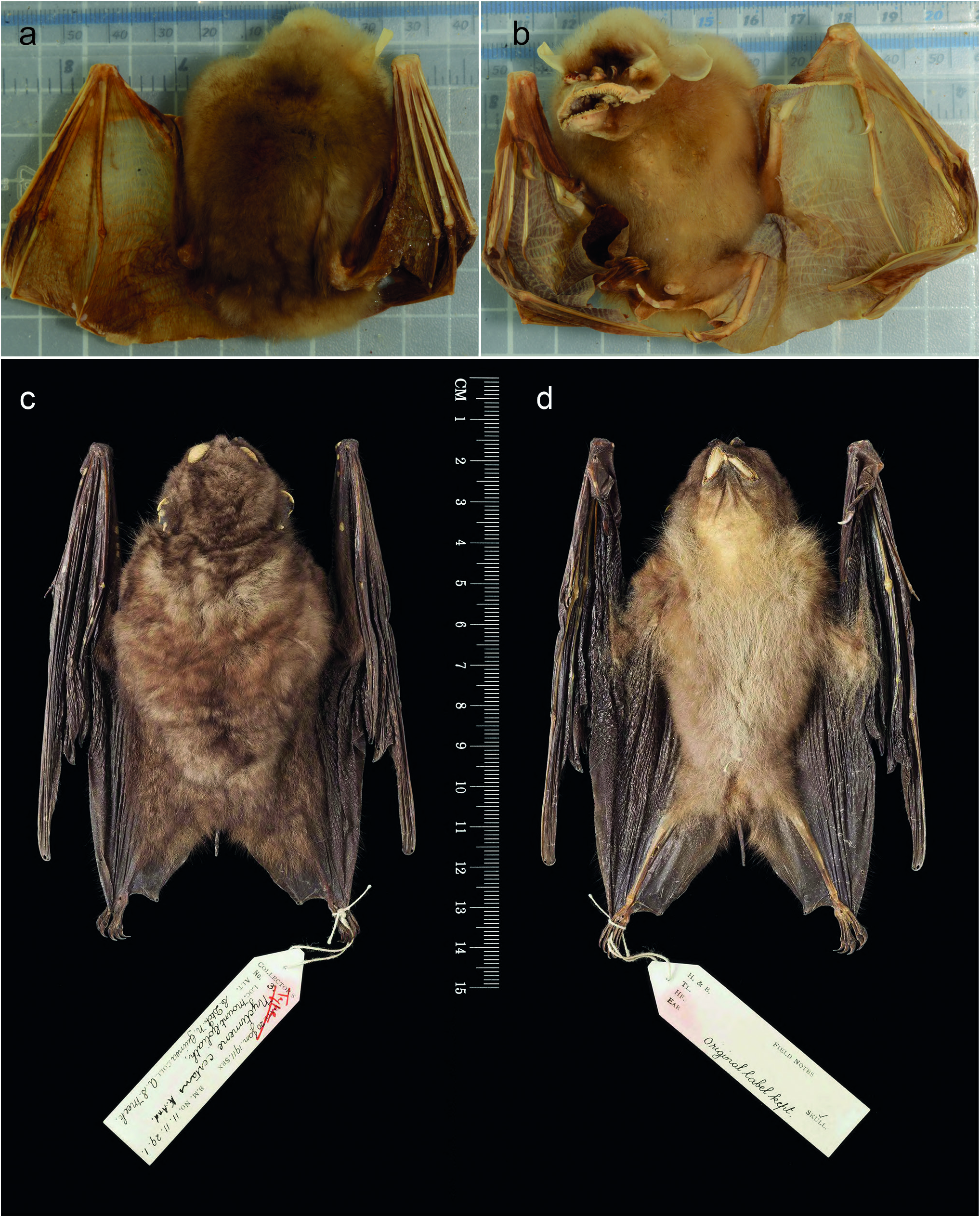
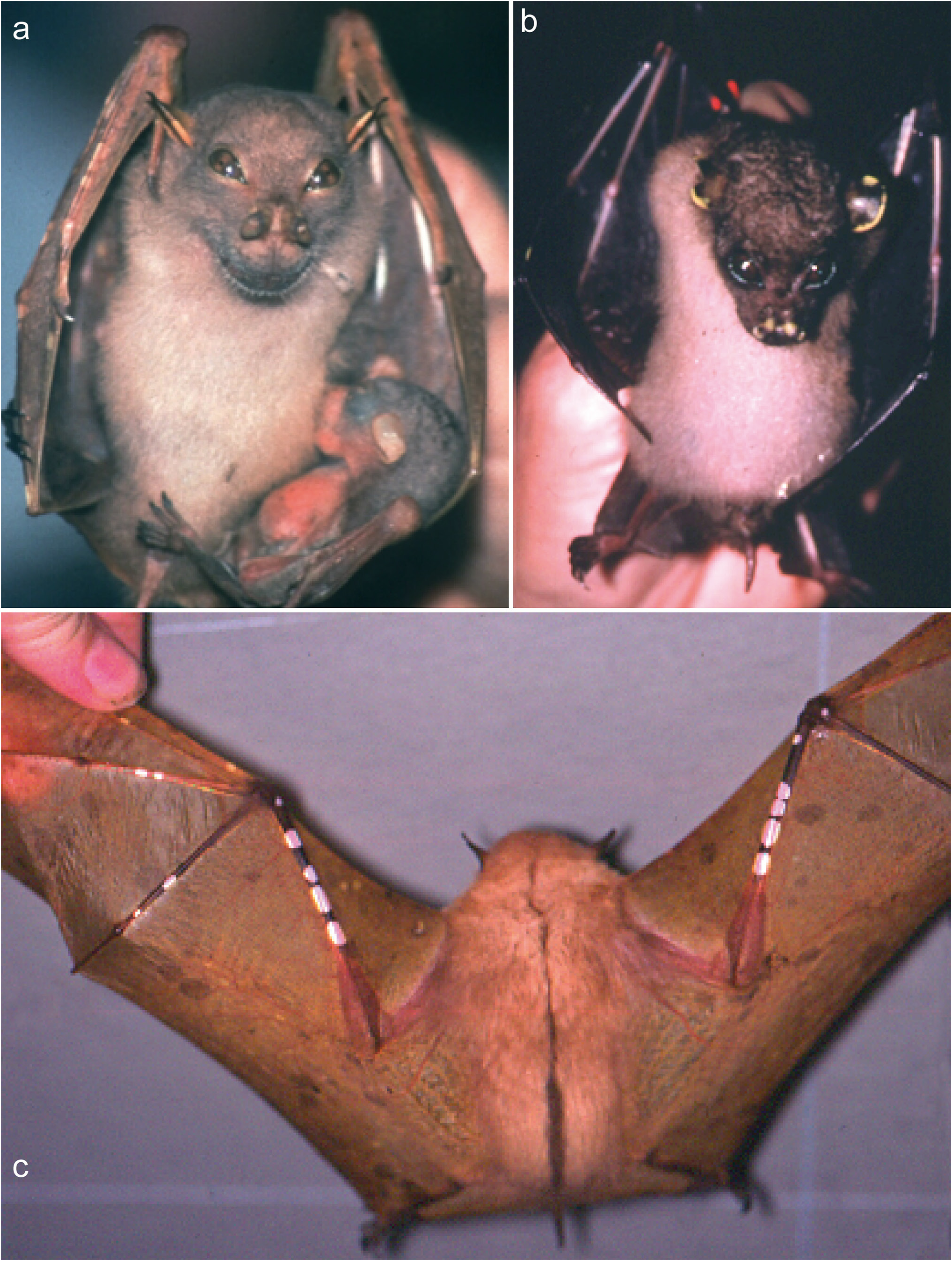
Nyctimene wrightae sp. nov. is difficult to identify in the field from sympatric species of Nyctimene , the key features are its brown fur, distinct dorsal stripe, long pointed ears, generally large size and its distinctive “smiling” facial expression ( Fig. 7 View Figure 7 ).
Description. The ears are similar in shape to N. a. papuanus , they are thickened on the leading edge, which is a feature more similar to N. certans and N. cyclotis (see Figs 5–7 View Figure 5 View Figure 6 View Figure 7 ). Ear length scales with body size in N. wrightae sp. nov. Fur colour of N. wrightae sp. nov. is variable but is typically brown on the dorsal, grey-brown around the face and the pale venter contrasts with the browner fawn colour of the dorsum ( Figs 6 View Figure 6 , 7 View Figure 7 ). The wings have variable white (sometimes yellow) and brown spotting typical of the subfamily. Large white or yellow spots are found more commonly on the large metacarpals of the wings ( Fig. 7 View Figure 7 ). Individuals vary greatly in the extent of spotting. Nyctimene wrightae sp. nov. shows no consistent pattern of sexual size dimorphism. Sexual dimorphism in colour is more pronounced in the reproductive season. The hair of the central thorax is whiter and brighter in males than females. Males have brighter, more defined fur colour especially on the ventral flanks where they have striking yellowish-orange patches, which develop during the mating season. Females are an overall fawn brown colour and are generally duller. Dr Deb Wright and I have occasionally caught yellow morphs from three locations in mainland New Guinea ( Fig. 7c View Figure 7 ). These morphs have yellow dorsal and ventral fur with brown wing spotting and white spotting on the forearm and wing digits.
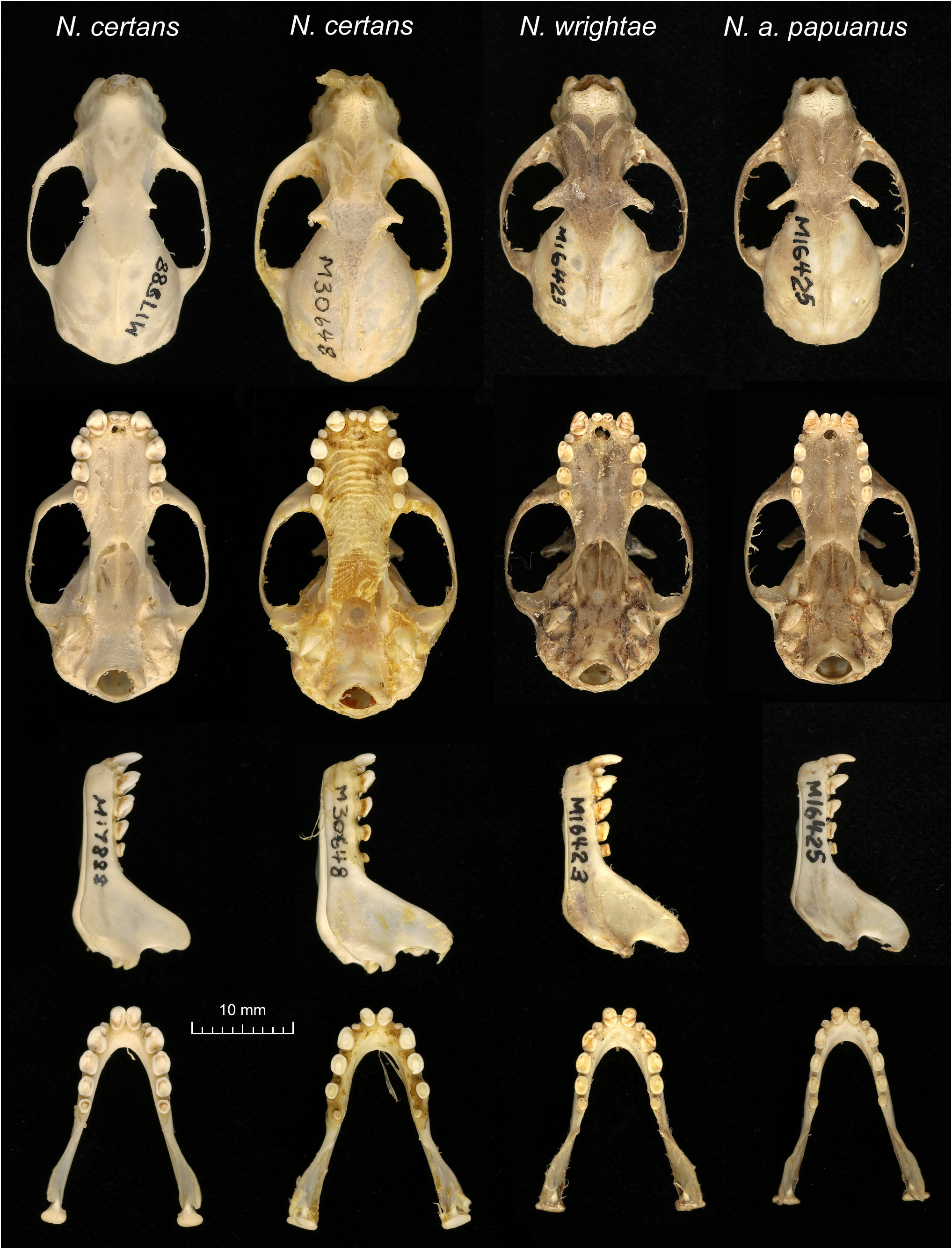
The palate length is relatively short compared to GSL. The rostrum is short, R-L averaging 3.62 mm (± 0.6 SD) and is deep dorsoventrally. Teeth are broad and cuspidate ( Fig. 4 View Figure 4 ).
Mean P 4 -P 4 (8.61± 0.7 mm) is equal to or greater than mean M 1 -M 1 (8.49± 0.6 mm) in N. wrightae sp. nov., unlike N. a. papuanus , in which M 1 -M 1 averages larger (7.92± 0.80 mm) than P 4 -P 4 (7.71± 0.76 mm). The upper premolars and molars are generally broad and semi-circular in outline in N. wrightae sp. nov. and almost as broad as long. This contrasts with the equivalent teeth in N. a. papuanus , which are less broad and are on average smaller and more rectangular in shape (longer in length than breadth); N. certans have squarer, broader teeth and relatively much broader cheek teeth (both upper and lower P 3 and P 4) than either N. a. papuanus , N. wrightae sp. nov. or N. cyclotis ; and N. cyclotis has premolars that are similar in shape (longer than broad) to N. a. papuanus but which are much bigger ( Fig. 4 View Figure 4 ). The brain case length of N. wrightae sp. nov. is shorter relative to skull length than N. certans , N. cyclotis , or N. a. papuanus ( Figs 2 View Figure 2 , 4 View Figure 4 ).
Etymology. With great pleasure I name this bat after Dr Debra Wright, Director for Wildlife Conservation Foundation, PNG 1996–2003, the co-founder of PNG Research Foundation and PNG Institute of Biological Research. She has tirelessly given her energy and considerable skills to capacity building and training Papuan New Guinean scientists. She provided logistic assistance for my team and I, helped me secure funding, provided access to facilities, and gave moral and intellectual support throughout my field-work. Her good humour, boundless energy, passion and generosity meant that despite spending many years together in remote places, we have formed a lifelong friendship. It was this cryptic bat species that started my whole journey with Nyctimene .
Common name. Happy tube-nosed fruit bat. Field assistants Toby Kamli and Rafael Kaneg, noticed the difference between this species and N. a. papuanus in the hand and called this species the hamamas bat, which in Tok Pisin means the happy bat, due to the wide “smile” characteristic of this species. I therefore propose the happy tube-nosed fruit bat as the common name, alluding to the “smile” that seems to result from the broad dental arcade and palate and very short rostrum, which are diagnostic characters of the species.
Remarks. Taxonomists have known for some time that there were aberrant specimens with affinities to N. a. papuanus or N. certans that did not fit described forms of Nyctimene . Tate (1942) noted an odd specimen from the Idenburg River, Papua that differed from the other 44 specimens of N. a. papuanus from the Fly River, PNG in the Archbold collections. Tate (1942, p. 721) states “The only observable difference … is the wider molars, but even so their width comes within the upper range given by Andersen in his table”. Likewise, McKean (1972) mentions a specimen from Lake Kutubu, Southern Highlands Prov., PNG (CM2316 from ANWC) that had skull and dentition similar to the description of the holotype of N. cyclotis , but a quite different pelage, being “short and brown rather than long, grey and mottled”. The specimen was rather large, with a FA of 62.1cm given by McKean. He was at pains to point out that the skull and skin were not mismatched. Flannery (1995b) identified the same Lake Kutubu specimen as N. cyclotis . I have not examined the latter specimen nor the one from Idenburg River discussed by Tate, but from the descriptions and the measurements provided by the latter authors it is highly likely that both specimens are representatives of N. wrightae sp. nov. Although not discussed by McKean, another slightly smaller individual that he lists as N. a. papuanus (ANWC 2317, FA = 58.5 mm), was caught from the same locality at Lake Kutubu on the same day. It would be interesting to confirm the identity of this specimen, to assess a possible instance of sympatry between N. a. papuanus and N. wrightae sp. nov.
The N. wrightae sp. nov. paratype BPBM28396 from Mt Missim, Morobe district was previously identified as N. certans by Peterson (1991) and illustrated in fig. 2 of that paper. The splayed P 3 are clearly evident in his illustration, and the dental arcade, while rounded, is not as extreme as in N. certans . The brain case is short in length compared to the GSL, characteristic of N. wrightae sp. nov.. The measurements of the 5th metacarpal and 5M-1P compared to the 2nd-2nd also distinguish this specimen from N. certans . Several N. certans (BPBM 28397–9, 28404, and 28410) were caught sympatrically with the N. wrightae sp. nov. paratype at Mt Missim. The other five BPBM paratype specimens listed above were individuals previously assigned to N. cyclotis by Peterson (1991) and Bonaccorso (1998).
NHMUK 1897.12.6.3 reassigned here as a paratype of N. wrightae sp. nov., is one of 15 specimens identified as N. papuanus (currently placed as N. a. papuanus ) by Andersen (1912a).
Nyctimene wrightae sp. nov. is restricted to mainland New Guinea. The extensive intraspecific variation found within N. wrightae is likely to have a geographic component, perhaps associated with latitude, altitude and niche sympatry.
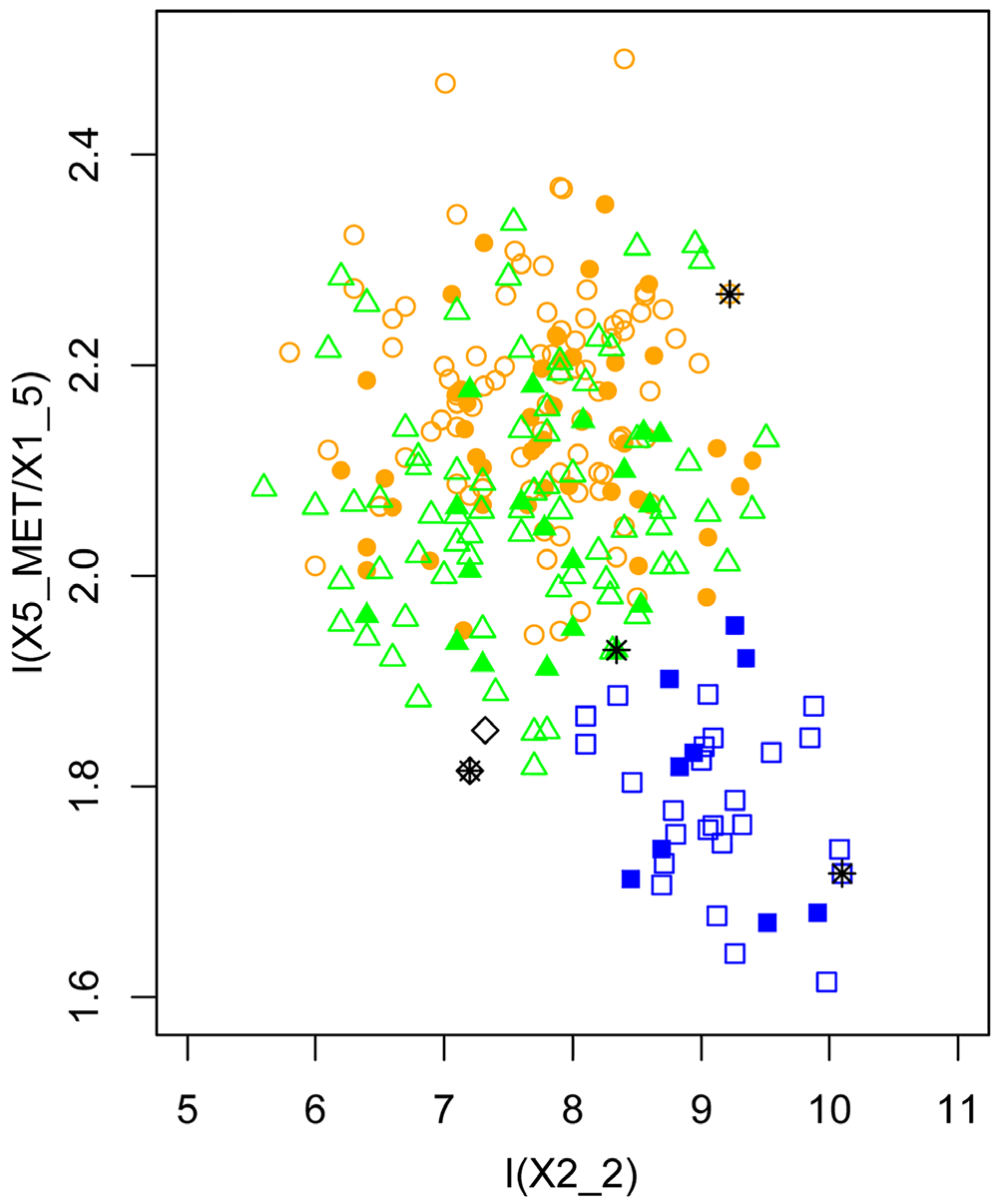
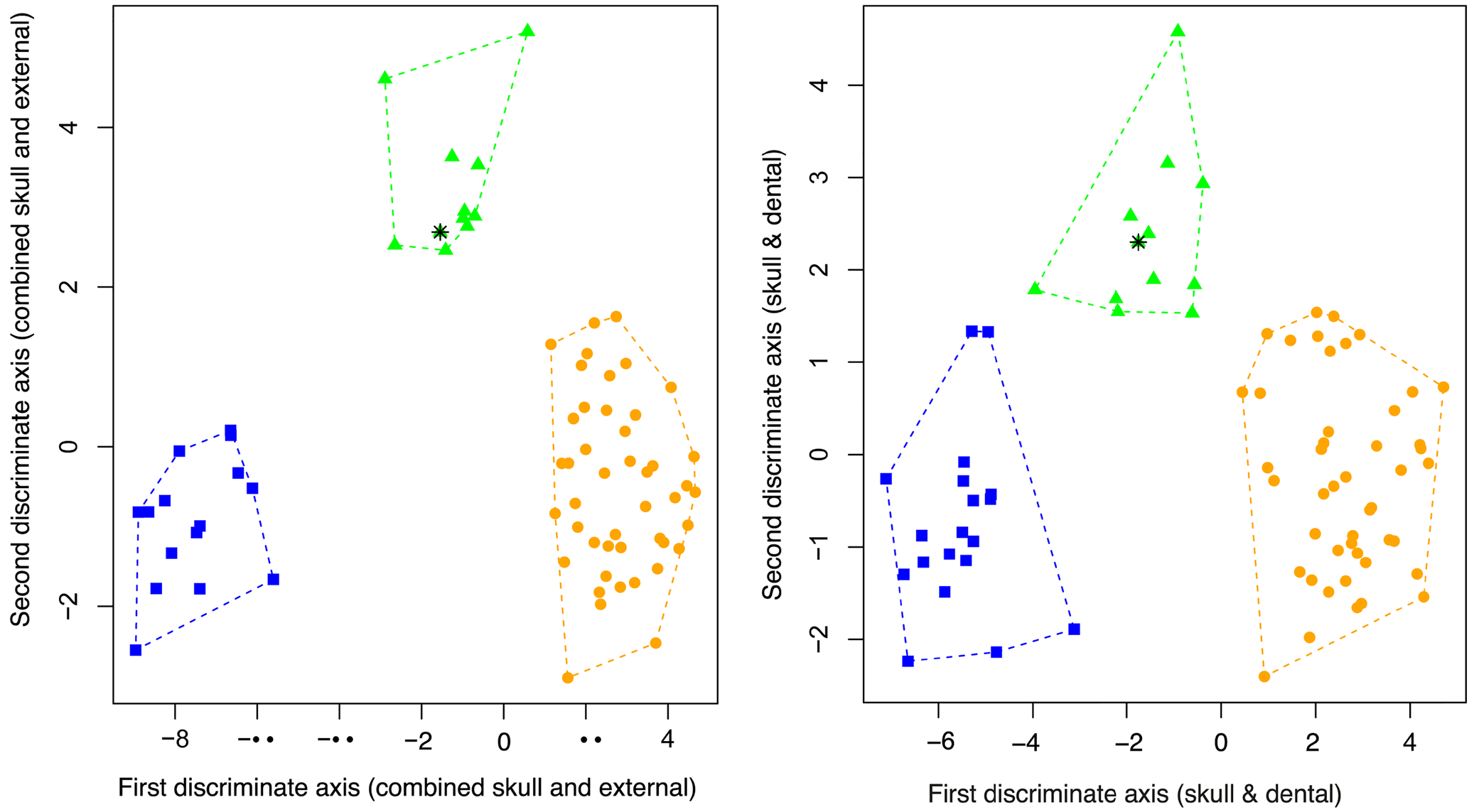
Reproductive biology. What little is known of the reproductive biology is from my field records combined with examination of museum specimens caught while pregnant. BPBM 28396 and WAM 47463 were caught in January and both had a large well-developed foetus, the latter foetus even had a well developed dorsal stripe. I caught pregnant females in Opio on 17 and 22 January (NRI 776–NRI 823) and as late as 14 February (NRI 888) during 2000. On two occasions during July, in the same area, I caught females with volant young that were of equal or larger body size of the mother and were still suckling, which suggests a long maternal care investment of about 6 months. In general the pattern therefore seems to be that pregnancy occurs from January to February, with females still lactating with volant young in July Morphometric analyses. Separation between the three species N. wrightae sp. nov., N. certans , and N. a. papuanus was achieved by using a LDFA with 14 variables (some composite) derived from 4 wing and 13 dental and skull measurements for 71 specimens ( Table 4, Fig. 8 View Figure 8 ). The predictive accuracy of the model at classifying specimens to the correct species was 100%, with a posterior probability of assignment> 99.9 for all specimens. The loadings on dimension one explained 0.9017 of the proportion of the trace and while influenced by all the variables the first dimension is most influenced by variables that reflect the breadth of the skull, (Zygo-B/GSL, Brain-W/Brain-H) and the breadth of the premolars (P 3 and P 4). LD2 explained only 0.0983 and was overwhelmingly influenced by the length of the zygomatic arch in proportion to the length of the skull (Zygo-L/GSL) and the breadth of the molar teeth (P 4, M 1 and M 2) ( Table 4). Tooth breadth was particularly useful for separating these species but as it is subject to tooth wear with age, species separation was therefore less clear when analyses were confined to skull and dental measurements ( Table 5, Fig. 9).
Species comparisons
Nyctimene cyclotis is similar to N. certans and to the exclusion of other Nyctimene from New Guinea, shares a grizzled and long woolly fur. Nyctimene a. papuanus and N. wrightae sp. nov. are in contrast generally brown taupe with short fur. The short rounded ears N. cyclotis is diagnostic of that species, in combination with the indistinct dorsal stripe in the lower portion of the dorsum and straight parallel post canine tooth row and small M 1 relative to P 4. Nyctimene cyclotis is distinguishable from N. a. papuanus on the basis of its larger cheek-teeth and broader dental arcade (esp. at P 3 -P 3) and smaller M 1 relative to P 4 ( Table 2; Fig. 3 View Figure 3 ). Andersen (1912a) noted that N. cyclotis had similar pelage to N. varius , while N. minutus and N. vizcaccia (unknown to Andersen) also have the similar grizzled dorsal fur and ill-defined dorsal stripes, but do not share other features in common with N. cyclotis or N. certans , such as ear shape or the shape and dimensions of the palate, dental arcade, or teeth. Genetic sequencing from N. cyclotis would enhance considerably the understanding of the species and placement of N. cyclotis in a phylogeny of the subfamily.
Nyctimene certans tends to be larger than N. cyclotis , N. wrightae sp. nov. and N. a. papuanus , as shown by its significantly larger FA than the latter species ( Table 2). The ears of N. certans are short relative to its overall body size, similar to N. cyclotis . Raw measurements of mean ear length were not significantly shorter in N. certans compared to N. a. papuanus ( Table 3). However, when taken as a proportion of body size (Ear/FA), ear length was significantly shorter compared to both N. a. papuanus (Welch t -test, n 1 = 30, n 2 = 131, t = -5.19, p = 7.546E- 06) and N. wrightae sp. nov. (Welch t -test, n 1 = 36, n 2 = 101, t = -5.21, p = 7.438E- 06). Ear shape, being round rather than tapered on the distal portion, is immediately diagnostic rather than actual length measurements, which can be very difficult to measure in the field on a live bat. Nyctimene certans have long ( 10–14 mm) hair reflective of their mountain lifestyle ( Fig. 5 View Figure 5 ).
Although N. certans is generally larger than N. cyclotis , the range for some characters overlap between these species and further samples of N. cyclotis might result in overlap in additional characters ( Table 2). The main differences between the holotypes of N. certans and N. cyclotis are palate shape, the broadness of the dental arcade, width of the palate and the broader check teeth of N. certans . The type description of N. certans describes the broader teeth than other sympatric species and a P 4 greater than 1.7 and M 1 -B greater than 1.4 mm, would also be indicative of this species ( Table 1, Fig. 10 View Figure 10 ).
Flannery (1995b) stated that N. certans and N. cyclotis occurred sympatrically in 5 locations and could be distinguished by different CBL, which he regarded as possibly the only useful diagnostic character to separate these species. He did not provide specimen registration numbers, but I think he was actually comparing N. wrightae sp. nov. and N. certans . Nyctimene wrightae sp. nov. is significantly shorter in GSL and CBL to N. certans , ( Tables 2, 3), confirming Flannery’s observation. GSL of N. wrightae sp. nov. is also significantly longer than N. a. papuanus ( Fig. 4 View Figure 4 ; Tables 2, 3). The posterior of the skull is missing in both specimens of N. cyclotis and therefore no comparison is possible at present for this species. GSL and CBL measurements overlap between medium sized Nyctimene species and although mean values might be significantly different, absolute measurements are not diagnostic.
Nyctimene wrightae sp. nov. is generally of smaller body size than both N. cyclotis and N. certans but usually larger than N. a. papuanus . Although there is extensive overlap in the FA ranges between all of the four latter species ( Table 2), mean FA of N. wrightae sp. nov. is significantly smaller than that of N. certans and significantly larger than N. a. papuanus ( Table 3). Ear shape of N. wrightae sp. nov. is similar to N. a. papuanus in that the ears taper distally in both species, in contrast to the short rounded ears of N. certans and N. cyclotis . The ears of N. wrightae sp. nov. are thickened on the leading edge, similar to N. certans and N. cyclotis , but unlike N. a. papuanus . Ear length is significantly longer in N. wrightae sp. nov. than in both N. a. papuanus and N. certans based on summary statistics ( Tables 2 and 3). However, ear length scales with body size in these three species and if ear length is expressed as a proportion of body size (EAR / FA), no statistically significant difference remains between N. wrightae and N. a. papuanus (Welch t -test, t = -0.3251, n 1 = 95, n 2 = 45, df = 225.358, p = 0.7454) but does remain between N. wrightae and N. certans (Welch t -test, t = 5.1416, n 1 = 95, n 2 = 36, df = 47.166, p = 5.163E- 06). Measurements of CBL as a proportion of GSL indicates that N. wrightae sp. nov. has relatively longer CBL than other small-bodied Nyctimene , with mean CBL/GSL equal to 84% in N. wrightae sp. nov. as opposed to 81% in both N. certans and N. a. papuanus ( Table 3, Fig. 4 View Figure 4 ).
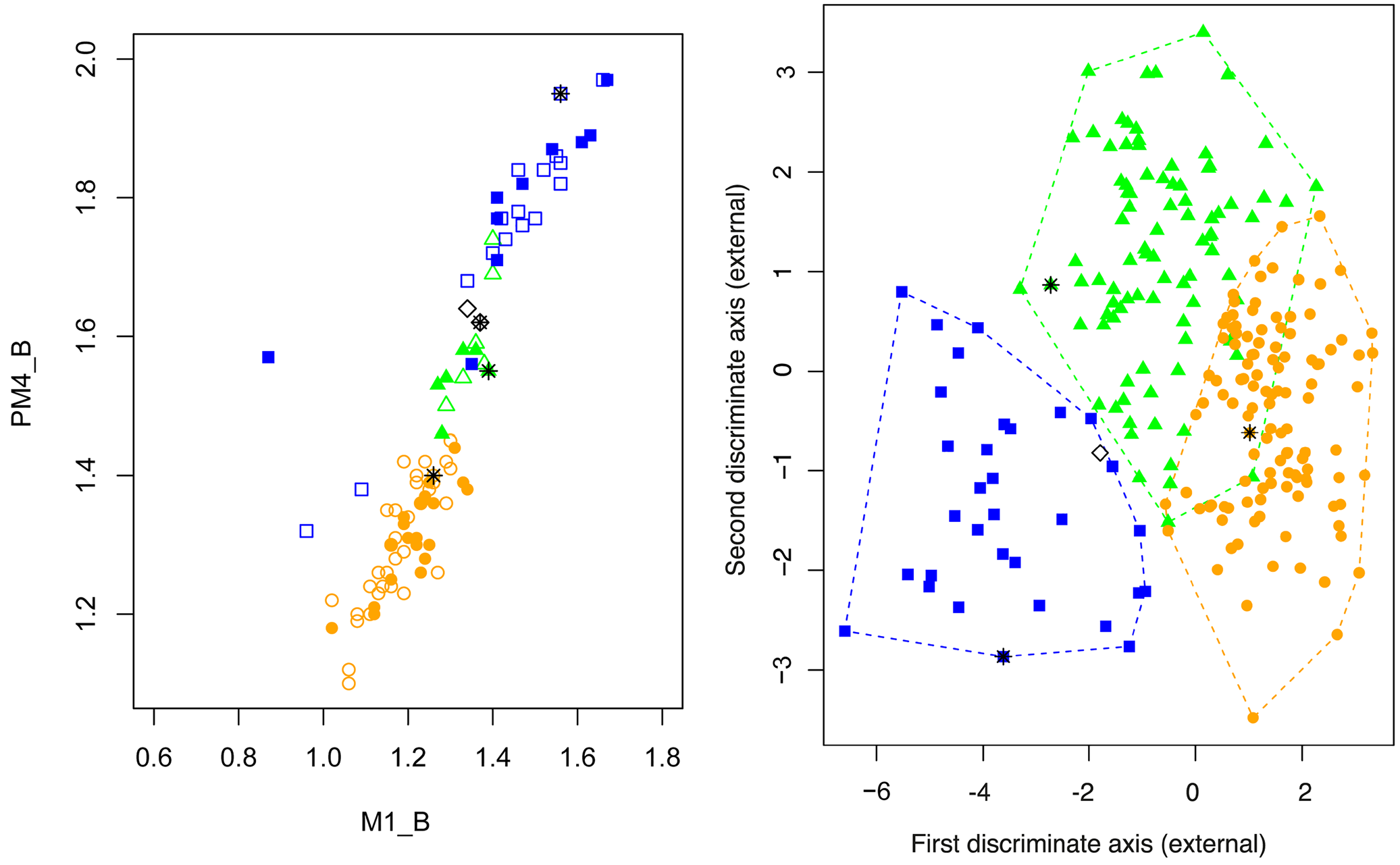
Morphometrically, N. wrightae sp. nov. and N. a. papuanus are difficult to separate on external variables alone and are therefore truly cryptic. Although ranges of wing measurements overlap ( Table 2), mean wing measurements of N. wrightae sp. nov. were significantly larger than those of N. a. papuanus for metacarpals and phalanges, see Table 2. In a LDFA based on 7 external wing elements (all of which could be measured in the field), some separation of specimens of N. wrightae sp. nov. from N. a. papuanus can be achieved, but there is considerable overlap ( Fig. 11) ( Table 6). No particular variable contributed greatly to the loadings ( Table 6). The LDFA assigned 228 out of the 245 individuals correctly to the pre-assigned species (93%), but the confidence in the classification was sometimes low. Even when identification was correct the confidence in the prediction from the model was not always high; the model correctly classified, 119 of the 120 N. a. papuanus but 16 had a posterior probability of <0.75; 28 of the 32 N. certans were corrected classified with 3 with low confidence; whereas 81 of the 93 N. wrightae sp. nov. were correctly assigned, but 10 with low confidence. Nyctimene wrightae sp. nov. was therefore the species most likely to be misclassified, and always identified incorrectly as a N. a. papuanus . Species separation was clearer in a LDFA using 14 skull and dental measurements that described palate shape, tooth breadth and skull length and breadth ( Table 5, Fig. 9). All specimens were assigned correctly to species with high accuracy (posterior probability> 98%). Nyctimene wrightae sp. nov. has a significantly broader skull (R-B, Zygo-B and Brain-W) relative to its size than N. a. papuanus and N. certans ( Table 3); and it was these skull breadth variables that contributed to the loadings that separated N. wrightae sp. nov. from the other two species. Nyctimene wrightae compared to N. a. papuanus has a relatively smaller M 1 -B compared to P 4 -B which gives an alpha character indication of species identification between the two very similar species. A bivariate plot of P 4 -B versus M 1 -B shows some separation between specimens of N. a. papuanus , N. wrightae sp. nov. and N. certans ( n = 98) ( Fig. 10 View Figure 10 ). Nyctimene a. papuanus and N. wrightae sp. nov. separate in my current dataset ( Fig. 10 View Figure 10 ). However, these criteria on their own are not sufficient for unambiguous species separation between all the species as there is a degree of overlap between N. certans , N. wrightae sp. nov. and N. cyclotis . These variables are however very useful in conjunction with other variables in a LDFA which contribute to the separation of the species ( Figs 8 View Figure 8 , 9). Measurements of these two teeth variables must be of adult specimens with “reasonable” levels of tooth wear; tooth breadth is greater in younger animals before they have worn their teeth and much reduced in older animals as they literally wear out their teeth. Where specimens fall on the plot is both dependent on the species identification and the age of the specimen (see Fig. 10 View Figure 10 ). Young adults of N. wrightae sp. nov. fall in the range of aging N. certans and older specimens of N. wrightae sp. nov. fall into the range of young adults of N. a. papuanus . In addition, reduction in tooth dimensions from excessive wear is so marked in some specimens of N. certans that four specimens fall outside of the strong colinear pattern shown across species in Fig. 10 View Figure 10 . In these four specimens, using P 4_ B or M 1 _ B measurements to inform a LDFA for identification would therefore not be recommended. The two specimens of N. cyclotis shown in Fig. 10 View Figure 10 indicate that their tooth dimensions conform to the linear size continuum of the small-bodied Nyctimene in New Guinea, being larger than N. a. papuanus and much smaller than the majority of N. certans . To increase both the predictive power and accuracy of diagnosis, a combined LDFA of skull and external measurements is recommended ( Fig. 8 View Figure 8 ).
I caution overconfidence in assigning specimens from only a few external measurements, as an LDFA model can predict the wrong species. NHMUK 1969.147 for example is a N. wrightae sp. nov. ( Fig. 6 View Figure 6 ) but on the 7 external body measurements chosen in Fig. 11 is predicted to be a N. a. papuanus . The predictive score for assignment is low using these external measurements, (p.p. <60% assigned to N. a. papuanus ), but by increasing the external measurements to 12 variables an LDFA predicted the specimen to be a N. wrightae sp. nov. (51%); increasing the number of measurements, while increasing the noise, increases the posterior probability (variables = 16, p.p. assignment to N. wrightae sp. nov. = 73%). The skull of NHMUK 1969.147 however is clearly a N. wrightae sp. nov. (p.p.> 98 %) and the combined analysis of external and skull measurements ( Table 4) assigned this specimen unequivocally to N. wrightae (99%). One explanation of the difficulty of assignment could be a mix up of the skin and skull. However, an alternative explanation is that N. wrightae sp. nov. and N. a. papuanus are phenotypically conservative and there could be many individuals that are difficult to distinguish from external morphometrics alone.
In summary, N. wrightae sp. nov. differs from N. certans and N. cyclotis in having shorter fur that is brown not greybrown grizzled, longer tapered ears, a clearly defined and longer dorsal stripe, a short rostrum, and premolars that are longer than broad but with a broader M 2 relative to P 4 ( Fig. 4 View Figure 4 ). The dorsal stripe on N. cyclotis and N. certans by contrast is very difficult to see as the long, grey-brown grizzled hair makes it difficult to distinguish. The dorsal stripe of N. wrightae sp. nov. tends to be thinner than in N. a. papuanus . Nyctimene wrightae sp. nov. is on average larger in all dimensions and has a broader palate (wider or sup-equal P 3 -P 3 compared to P 1 -M 1 length) than N. a. papuanus (narrower P 3 -P 3 compared to P 1 -M 1 length). This gives the species a robust facial appearance and the difference in dorsal stripe, along with differences in the wing phalanges, provide characters that for some individuals will be indicative of their species using external features ( Fig. 11), however a more definitive diagnosis requires an examination of the dental arcade and cheek teeth ( Figs 8 View Figure 8 , 9).
| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |