
Varanus dalubhasa, Welton, Luke J., Travers, Scott L., Siler, Cameron D. & Brown, Rafe M., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3881.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:62DB7048-70F2-4CB5-8C98-D7BCE48F4FC2 |
|
DOI |
https://doi.org/10.5281/zenodo.6139289 |
|
persistent identifier |
https://treatment.plazi.org/id/6D728781-FFC5-3A0A-C4C7-FC4DFBE1A081 |
|
treatment provided by |
Plazi |
|
scientific name |
Varanus dalubhasa |
| status |
sp. nov. |
Varanus dalubhasa sp. nov.
Figs. 3 View FIGURE 3 , 5–8 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8
Holotype. PNM 9796 (formerly University of Kansas Biodiversity Institute [KU] 306603; Field No. CWL 521), adult male, collected by Charles W. Linkem and CDS, 0 8 July 2006, 18 m above sea level (14.03202, 122.34143; WGS-84), Barangay Madlangdungan, Municipality of Calauag, Quezon Province, Luzon Island, Philippines.
Paratopotypes. KU 305155 ( CDS Field No. 2202), adult male; PNM 9797 (formerly KU 306601; CWL Field No. 440), juvenile; KU 306602 (CWL Field No. 520), adult male.
Paratypes. KU 308216 ( CDS Field No. 2298), juvenile, collected 22 February 2007, Barangay Buenavista, Municipality of Bato, Catanduanes Province, Catanduanes Island, Philippines; KU 313880 (RMB Field No. 9910), adult male, collected 0 1 July 2008, Barangay Tulay Na Lupa, Municipality of Labo, Camarines Norte Province, Luzon Island; KU 326702 and 326703 (LJW Field No. 0 0 75, 0077), adult males, salvaged 3 August 2009, Polillo Island (exact locality information unknown).
Other Material. See Specimens Examined Section.
Etymology. The specific epithet, dalubhasa , is derived from the Tagolog word “ dalubhasa ” meaning a person who has authoritative and comprehensive knowledge of a particular area, or a skilled expert in a particular subject. We choose this term in honor of Vicente “Enteng” Yngente of Polillo Island, whose extensive knowledge of natural history and ecology of Philippine reptiles (particularly monitor lizards) has been instrumental to our research and conservation work. Suggested common name: Enteng’s Monitor Lizard.
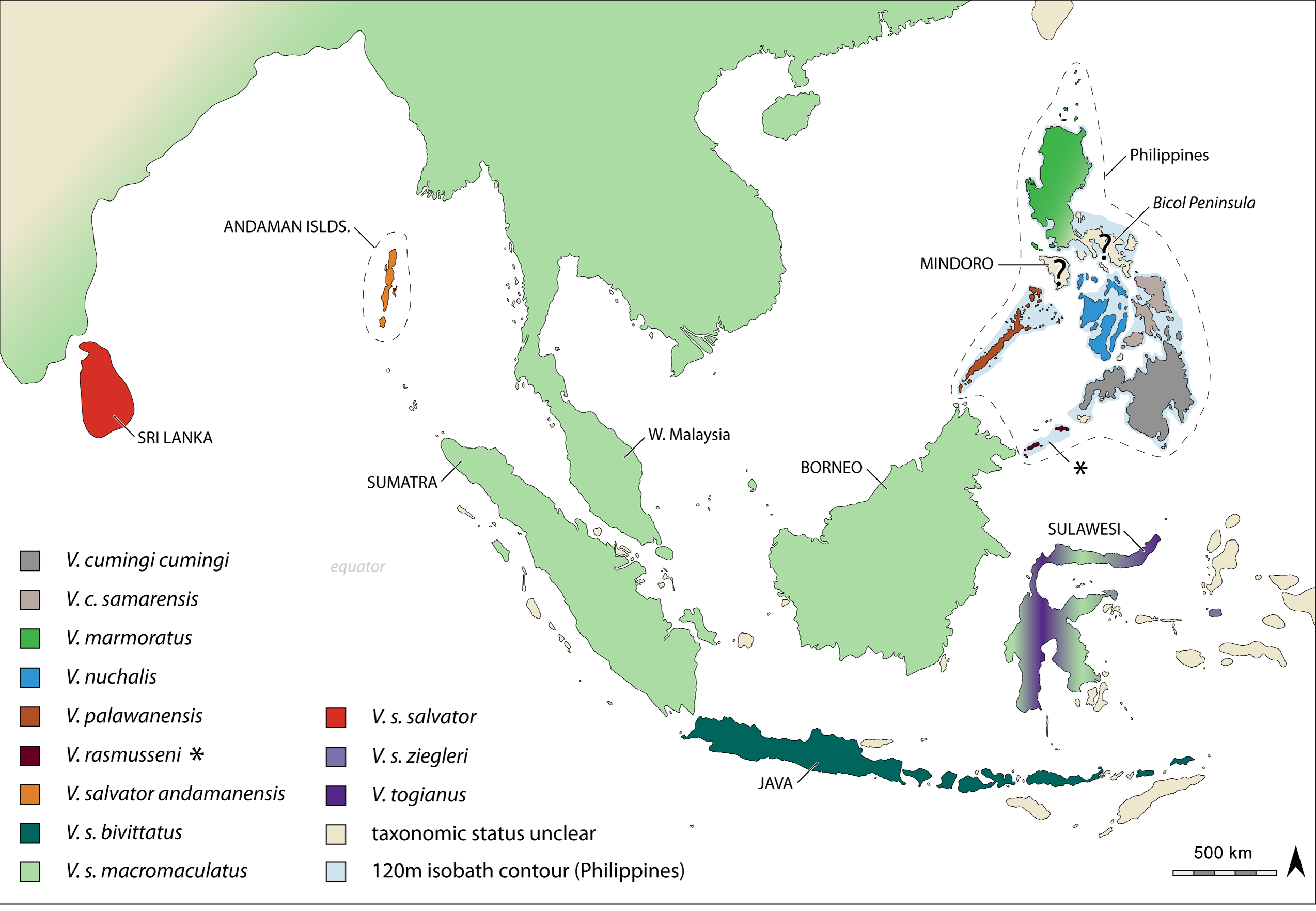
Diagnosis. Varanus dalubhasa can be distinguished from congeners by (1) small, dark speckling and variable transverse banding through the gular region ( Fig. 3 View FIGURE 3 ); (2) number of scales around the tail at 1/3 distance from the base; (3) number of gular scales; (4) number of dorsal scales in nuchal region; (5) total number of dorsal scales; (6) number of scales around the neck anterior to the gular fold; and (7) phylogenetic placement sister to V. nuchalis . Additionally, this distinct lineage is biogeographically circumscribed in the Bicol Peninsula faunal sub-region, a distinct geological component of greater Luzon Island which remained inuslar until 3 Ma ( Hall 2002).
Comparisons. The new species in phenotypically nearly indistinguishable from Varanus marmoratus , but can generally be diagnosed by the presence of small, dark speckling in the gular region and faint anterior transverse bands (versus speckling and distinct anterior transverse bands; Fig. 3 View FIGURE 3 ), and its allopatric distribution in the Bicol faunal sub-region (versus the remaining portions of Luzon and Lubang islands, and the Batanes and Babuyan island groups). Additionally, although V. dalubhasa is phenotypically similar to V. marmoratus , available data suggest that it is not most closely related to this species ( Fig. 2 View FIGURE 2 ).
Varanus dalubhasa can be conveniently distinguished from its allopatric sister taxon, V. nuchalis (distributed in the West Visayan islands of Negros, Panay, Guimaras, Masbate, and the Romblon Province islands of Sibuyan, Tablas, and Romblon; Fig. 1 View FIGURE 1 , 2 View FIGURE 2 ), by the presence of more scales around the tail at 1/3 distance from the base (mean = 53 ± 4, versus 46 ± 3), fewer gular scales (mean = 80 ± 3, versus 73 ± 3), and more dorsal scales from the tympanum to the gular fold (mean = 27 ± 2, versus 23 ± 2). Of the remaining, geographically proximate species of the V. salvator Complex, this new species can be distinguished from V. palawanensis by having fewer dorsal scales from the tympanum to the gular fold (mean = 27 ± 2, versus 32 ± 3), fewer dorsal scales from the gular fold to the hind limb insertion (mean = 84 ± 5, versus 96 ± 3), fewer total dorsals scales longitudinally (mean = 111 ± 5, versus 128 ± 3), and fewer scales around the neck anterior to the gular fold (mean = 78 ± 3, versus 87 ± 4). Additionally, V. dalubhasa can be distinguished from the allopatric Mindoro lineage by having variable dark speckling and transverse bands in the gular region (versus distinct spotting; Fig. 3 View FIGURE 3 ). Lastly, V. dalubhasa is comprised of entirely distinct haplotypes or haplotype networks, relative to all other members of the V. salvator Complex (see Welton et al., 2010a). We have constrained our morphological analyses to the geographically most proximate taxa due to previous studies (Koch et al. 2007, 2010b) demonstrating their distinctiveness relative to the remaining diversity withing the V. salvator Complex.
Description of holotype. An adult male ( Fig. 5 View FIGURE 5 ), hemipenes everted; SVL 505.0 mm; tail 657 mm; head relatively slender, length 89.1 mm, width 51.0 mm, and depth at eye 32.0 mm; snout length 47.6 mm, 53.4% head length; snout rounded anteriorly; narial openings 5.4 mm, ovular, posteriorly elevated, encircled by 9L/9R small polygonal scales; snout elevated sharply just anterior to narial region, then continuing gradually to ocular region; narial region not distinctly elevated above horizon of snout; cranial table squarish, only slightly wider than long.
Head scales ovular to polygonal ( Fig. 5 View FIGURE 5 ), heterogeneous in size with the smallest occurring dorso-laterally between ocular opening and tympanum, and largest ocurring dorso-medially from tip of snout to conspicuous parietal; supraoculars 6L/6R, subrectangular and elongated transversely; supralabials 59, slightly larger than lateral head scales; infralabials 61, smaller than both supralabials and nearest rows of gular scales; both supra- and infralabials increasing in size anteriorly; nuchals large, circular, increasing in size to forelimb insertion along both dorsal and lateral surfaces; lateral nuchals less than half the size of dorsals; scales immediately posterior to cranial table quite small, granular; sutures between scales slightly larger in nuchal region than on head; nuchals 31, in semi-regular transverse rows from posterior margin of tympanum to forelimb insertion; dorsal trunk scales arranged in 88 semi-regular transverse rows from fore- to hindlimb insertions, ovular, slightly smaller than nuchals, gradually decreasing in size to hindlimb insertion as well as laterally, with smallest scales occuring laterally adjecent to limb insertions; midbody scales 138; axilla–groin distance 245 mm; dorsal limb scales ovular, decreasing in size distally to digits; fore- and hindlimbs 92 and 107 mm, respectively, 18.2% and 21.2% snout–vent length, 32.6 and 53.9 mm diameter at insertions; limb scales present in semiregular rows; scales of manus and pes smaller and more irregular in shape, ranging from circular to nearly rectangular dorsally; supradigitals rectangular dorsally, ovular laterally, with terminal scale nearly twice as large as others; digits terminate in long, slender recurved claws; caudals ovular and granular at insertion, becoming rectangular posteriorly; dorsal keel composed of paired, posteriorly elevated scales commenses 65 mm posterior to hindlimb insertion, and continues to tail terminus with paired scales gradually decreasing in size; caudal scales moderately keeled, in regular transverse and longitudinal rows laterally, decreasing in size posteriorly.
Ventrals less variable than dorsals; gular scales generally rectangular anteriorly, becoming more ovular towards gular fold; smallest scales medially in anterior third of gular region, with largest scales just anterior to gular fold but followed posteriorly by three transverse rows of smaller, more granular gulars; scales around the neck anterior to the gular fold 78, mid-gular 87; gular scales in 85 irregular rows from tip of snout to gular fold; ventrals of fore- and hindlimbs circular to polygonal, with those of forelimbs slightly smaller than those of hindlimbs; precloacals homogenous in size and shape with those of hindlimbs; ventral trunk scales from gular fold to hindlimb insertion 85, ovular anteriorly, becoming rectangular posteriorly, and present in semi-regular rows; scales decreasing in size laterally and at hindlimb insertions; ventral caudal scales rectangular, smallest just posterior to cloacal opening; scale surface slightly rounded anteriorly, becoming sharply keeled and more longitudinally elongate posteriorly.
Right and left hemipenes partially everted; maximum length 56.2 mm, with proximal cylindrical section extending 26.2 mm; bifurcation resulting in primary cylindrical apical portion with external flounces, and more diminutive portion resembling an “ear” or reduced fold; flounces present in irregular rows, totalling 15 anterior to sulcus spermaticus, 6 posteriorly; lacking notable external appearance of ossified hemibaculum.
Measurements of holotype (in mm). SVL 505; tail length 657; head length 89.1, width 51.0, depth at eye 32.0; eye–naris distance 33.7; naris–snout distance 14.9; rictus–rictus scales 60; scales around tail base 103; scales around tail 1/3 distance form base 59; mid-body scales 138; gular scales 85; ventrals from gular fold to hind limb insertion 85; total ventral scales 170; dorsals from tympanum to gular fold 31; dorsals from gular fold to hind limb insertion 88; total dorsals 119; supralabials 59; scales around the neck anterior to gular fold 78; and enlarged supraoculars 6L/6R.
Coloration of holotype in preservative. Dorsal surfaces of head, body, and limbs black; aggregation of yellow-gold scales present dorsoanteriorly to narial openings, along canthal ridge just anterior to oculars, and irregularly dispersed throughout cranial table; trunk with seven transverse rows comprised of yellow-gold, indistinct ocelli; tail with nine, slightly more distinct yellow-gold bands, becoming more diffuse or speckled laterally; forelimbs with irregularly distributed yellow-gold scales, density increasing distally and along lateral surfaces; hindlimbs with irregular agreggations of ocelli consisting of only two or three yellow-gold scales ( Fig. 5 View FIGURE 5 ).
Lateral coloration of head consisting of partial, near vertical yellow-gold bands at the tip of snout, posterior margin of narial opening, and two between the narial opening and the ocular; posterior band with ventral widening oriented more posteriorly; post-orbitally, a yellow-gold stipe extends to tympanum, and a yellow-gold spot consisting of four or five scales appears ventral to the anterior margin of the stripe; lateral coloration of the nuchal region consists of 2L/3R faint spots directly posterior to the tympanum; ventral margins of nuchal region with yellow-gold variable demarcation, extending more dorsally along the posterior margin and appearing to wrap around the dorsal margin of the fore-limb insertions; trunk with variable yellow-gold spotting directly posterior to forelimbs, fading to solid black posteriorly ( Fig. 5 View FIGURE 5 ).
Ventral coloration predominately yellow-gold throughout; gular region with irregular spots, faint anteriorly but becoming prominent and dark along lateral margins just anterior to gular fold; scapular region with two irregular black bands; trunk with irregular black bands and spots anteriorly, becoming more broken medially and posteriorly, appearing as laterally elongated yellow-gold ovals; forelimbs with faint, irregular dark mottling; hindlimbs with near uniform venter medially, with dark mottling along lateral margins; manus and pes dark yellow-gray; venter of tail appearing anteriorly much like that of the trunk, with yellow-gold laterally elongated ovals; posterior portions of tail with more regular alternation of dark and yellow-gold bands ( Fig. 5 View FIGURE 5 ).
Variation. Our small type series exhibits some color variation, but specimens are generally phenotypically similar. The holotype (KU 306603) and three paratypes (KU 305155, 306602, and 308126) have lighter interstitial coloration between the dorsal scales, yielding an overall lighter appearance than other specimens. These three specimens also exhibit much more diffuse yellow gold coloration on limbs and tail, while two specimens (KU 313880, 326702) exhibit much more distinct yellow-gold spots or ocelli throughout. Additionally, the yellow-gold coloration in the lateral portions of the nuchal region, among all specimens, and on either side of a single specimen, are present as either a single longitudinal stripe extending posteriorly from the posterior margin of the tympanum or as a series of two or three longitudinally elongated spots or blotches. Two specimens (KU 305155, 306602) exhibit two relatively well-defined yellow crossbands on the snout, with the former’s being even more pronounced than the latter, and with this specimen (KU 305155) also exhibiting a much larger proportion of yellow coloration in the supraocular and temporal regions. In contrast, two specimens (KU 308216, 313880) exhibit nearly uniform, dark dorsal head coloration, with the former being completely devoid of yellow-gold accents with the exception of the pineal scale.
Ventral coloration is only slightly more variable than that of the dorsum. All specimens exhibit variable speckling in the gular region, gernerally consisting of 1–4 black scales. In the holotype (KU 306603), this speckling is much more reduced. In one specimen (KU 305155) anterior speckling in the gular region gives way to 2 broken transverse bands, and somewhat reduced speckling through the remainder of the gular region. Three specimens (KU 306602, 308216, and 326702) exhibit four, more well-defined dark transverse bands in the anterior portion of the gular regions, with speckling throughout the remainder of the gular region being relatively dense. A single specimen (KU 313880) exhibits nearly complete dark transverse bands throughout the gular region, with less prominent bands present anteriorly. Ventral trunk coloration is nearly uniform among specimens, with the only notable variation present in two individuals (KU 305155, 326702), the first of which exhibits reduced dark coloration extending medially onto the abdomen, and the second of which with darker, more distinct coloration extending medially onto the abdomen. Ventral tail coloration lacks any notable variation among specimens (see Figures 6 View FIGURE 6 and 7 View FIGURE 7 for further information on color pattern variation).
Coloration in life. Based on the only available image, PNM 9797 (formerly KU 306601; Fig. 8 View FIGURE 8 A), and field notes of the authors. Dorsal ground coloration black to dark gray, with consipcuous transverse cross bands of yellow-gold ocelli from the posterior margin of the nuchal region through the anterior third of the tail. Trunk ocelli larger with a darkened, ground-colored center. Additionally, dorsal trunk with faded reticulate pattern, owing to light brown interstitial coloration. Lateral margins of head and nuchal region with conspicuous yellow-gold stripe, extending from the the posterior margin of the ocular, through the tympanum, and midway into the nuchal region. Yellow-gold ocelli brightest at lateral margins of trunk and adjacent to hindlimb and tail insertions.
Dorsal limb surfaces colored similarly to body, but with ocelli irregularly arranged and significantly smaller, lacking any dark-colored center. Dorsal ground color of limbs bright yellow, with irregular dark pigmentation.
Ventral coloration bright yellow-gold, with scattered black pigmentation and with ocelli of ventro-lateral surfaces irregularly arranged and significantly smaller, lacking any dark-colored center ( Fig. 8 View FIGURE 8 A).
Ecology, Distribution, and Natural History. As with all members of the Varanus salvator Complex, V. dalubhasa should be considered a habitat generalist, often thriving in a range of habitats from primary forest to more urbanized areas. While no detailed studies have been undertaken on this taxon specifically, the findings of Gaulke (in Luxmoore & Groombridge 1989, 1992) are likely relevant. This taxon is most likely to be found in mangrove and riparian habitats (as their common name might suggest). Additionally, most water monitors seek out shelters in the form of rock crevices or even trees, most often near water sources. In more urban areas, human habitation may be used in lieu of more “natural” shelters ( Gaulke 1992).
Not surprisingly, Varanus dalubhasa has been found throughout the Bicol Peninsula and faunal sub-region, including genetically confirmed specimen identifications from Polillo and Catanduanes islands ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ). This is an area with significant agricultural (rice fields) and coastal environments, and is additionally subdivided by a number of moderately sized river drainages. The northern extent of distribution for this taxon, and potential contact zone with V. marmoratus requires further study.
Although no dietary preferences have been observed for this taxon, one can reasonably assume that its diet consists primarily of invertebrates and smaller vertebrates (fish, rodents, frogs, snakes). Additionally, this species is attracted to a number of carion items, with a particular affinity for fish (LJW pers. obs.).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.