
Isoperla vevcianensis Ikonomov, 1980
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5082.6.2 |
|
publication LSID |
lsid:zoobank.org:pub:2B45012F-EE84-4BF0-8DC8-25A03AD379EA |
|
DOI |
https://doi.org/10.5281/zenodo.5797608 |
|
persistent identifier |
https://treatment.plazi.org/id/6A554E4F-FFF0-D73E-FF0A-DA48FD94F971 |
|
treatment provided by |
Plazi |
|
scientific name |
Isoperla vevcianensis Ikonomov, 1980 |
| status |
|
Isoperla vevcianensis Ikonomov, 1980 View in CoL
( Figs. 1–27 View FIGURES 1–4 View FIGURES 5–8 View FIGURES 9–14 View FIGURES 15–20 View FIGURES 21–23 View FIGURE 24 View FIGURES 25–27 )
Isoperla vevcianensis Ikonomov, 1980 View in CoL — Ikonomov 1980: 26. (original description of the male, female and larva); Murányi 2011: 21 View Cited Treatment . (redescription of the male).
Type locality: North Macedonia, small streams above Vevchani , at elevations 1100, 1300 and 1450 m (Jablanica Mts) .
N ew records: NORTH MACEDONIA: Southwestern region , Jablanica Mts, Labuništa, open brook W of the city, 1905 m, N 41° 16.069’ E 20° 31.242’, 26.vi.2014, leg. Péter Juhász, Tibor Kovács, Dávid Murányi: 5♂ 1♀, 2 larvae ( HNHM: PLP 4840 View Materials ), GoogleMaps 7♂ 5♀, 1 larva ( MM) GoogleMaps .
Diagnosis. Both adult and larva with large head. Male medial penial armature with upper part trapezoid, lower part rounded to rounded triangular, lateral armatures lacking. Egg oval, with marked ornamentation of FCIs, collar with slightly flanged rim. Larva with pronotum half as long as wide, lacinia trapezoid shaped, paraproct blunt with widely rounded apex, cercus lacks dorsal hair fringe.
Description of the larva: Body length of the mature larva: 16.0–18.0 mm (n=3). General colour brown, with indistinct paler markings on the dorsum and pale ventral aspect ( Figs. 1, 3–4 View FIGURES 1–4 ). Pilosity usual for the genus, consists of fine black clothing hairs and acute marginal spines. Occipital row dense, medially interrupted only at the occipital suture ( Figs. 1 View FIGURES 1–4 , 9–10 View FIGURES 9–14 ); pronotal marginal fringe distinct, longest spines are as long as one tenth of pronotal width ( Figs. 1 View FIGURES 1–4 , 12 View FIGURES 9–14 ); posterior tergal fringe with acute spines nearly half as long as tergal length on tergum 5 ( Figs. 3 View FIGURES 1–4 , 13 View FIGURES 9–14 ); cercal apical fringes dense but short and acute, on medial cercomeres most are shorter than one sixth of the length of segments but one or two spines of the fringe reach about one third of the length ( Fig. 14 View FIGURES 9–14 ); dorsal hair fringe lacking from the entire cercus ( Figs. 7 View FIGURES 5–8 , 14 View FIGURES 9–14 ). Head mostly brown and lacks interocellar spot, yellow markings limited to a small spot on the occipital suture, tentorial callosities and M-line; occipital rugosities and lateral occipital spots obscure; eyes small, the head is rather wide and frontal area slightly elongated ( Fig. 1 View FIGURES 1–4 ). Scape and apical half of pedicel brown, following antennomeres paler; palpi and mouthparts light brown, with dark edges on the maxilla and mandible ( Fig. 11 View FIGURES 9–14 ). Lacinia bidentate, wide trapezoid shaped; subapical tooth more than half of the length of apical tooth; submarginal row with one A seta interrupted from three B setae, the last one is being much shorter than previous two; one thin marginal seta (TMS) and one dorsal seta (DS) present; 9–11 marginal setae (C) initially widely spaced, last few shorter and closer, difficult to differentiate from dorsal surface setae; 12–16 ventral surface setae (D) starting from submarginal setae B, ending posteriorly at approximately four fifth of the inner lacinia margin length; dorsal surface setae (DSS) continue from last marginal seta (C) as a single submarginal row along inner-lateral margin, ending before posteriormost ventral surface setae ( Figs. 15, 17, 19 View FIGURES 15–20 ). Galea with 6–10 surface setae and 3–4 apical setae, basally slightly swollen, reaching base of apical tooth ( Figs. 15, 17, 19 View FIGURES 15–20 ). Mandible with three molars and three incisor dens, molar brush dense and long ( Figs. 16, 18, 20 View FIGURES 15–20 ). Pronotum short and slightly narrower than head, length half of width; brown with indistinct paler rugosities ( Fig. 1 View FIGURES 1–4 ). Mesonotum and metanotum mostly brown with indistinct pale brown pattern, wingpads paler; outer margin of wingpads nearly straight ( Fig. 1 View FIGURES 1–4 ). Thoracic sternites entirely pale, furcasternites and furcal pits large and slightly darker ( Fig. 4 View FIGURES 1–4 ). Legs entirely brown, tarsal claw lengths equal ( Figs. 1, 3 View FIGURES 1–4 ). Abdomen divided laterally by pleura in the first two segments; abdominal terga brown with moderately distinct, paired oval pale patch medially ( Figs. 3 View FIGURES 1–4 , 13 View FIGURES 9–14 ). Ventral surface of abdomen pale, the distal segments slightly darker. Paraprocts brown, large and blunt, apex widely rounded ( Figs. 5, 7 View FIGURES 5–8 ). Cercus light brown to brown, cercomeres cylindrical; medial cercomeres twice as long as wide ( Fig. 14 View FIGURES 9–14 ).
Updated characters of the male: Paraprocts dark brown, blunt, wide and slightly recurved in dorsal view (Fig. 63 in Murányi (2011)). In caudal view, the base is wide and bears only a very small membranous portion; apex bluntly triangular, inner edge slightly sinuous ( Fig. 22 View FIGURES 21–23 ).
Penis ( Fig. 21 View FIGURES 21–23 ): Divided into four lobes and a tubular basal section when extruded, lateral lobes poorly developed. The medial penial armature is divided into an upper and a lower part, lateral penial armatures lacking. Lower part of the medial armature located on the ventral lobe, upper part is on the medial lobe between the vestigial lateral lobes. The upper part of medial armature trapezoidal, tapered towards basal section; length 350 μm, width 200 μm. The lower part of medial armature is rounded triangular or rounded, length 200 μm, width 220 μm. The scales are spike-like, all of the same width, but those on the lower part are less dense, and generally shorter; width 15 μm, length 30–100 μm. The ventral lobe is small and weakly separated from the basal section, mostly aspinulate besides bearing the lower part of medial armature; spike-like coloured scales of the armature gradually turn into colourless triangular scales basolaterally, triangular scales continued on the basal section. The medial lobe is large and spherical, bearing the upper part of medial armature; upper edge of the armature transitioning into colourless triangular scales, then rapidly transitioning into hydra-like scales that are covering a wide area on the ventrobasal surface, becoming denser towards margins of the scale field; apical area aspinulate, lateral margin connecting small lateral lobes with small hydra-like scales. Lateral lobe is poorly developed and hardly delimited from medial lobe; mostly smooth, but with lateral band of small hydra-like scales, and a few sensillae. The basal section is completely covered by triangular scales.
Egg: Chorion brown; length 0.28–0.35 mm, width 0.22–0.26 mm (n=8). Shape oval, opercular end usually depressed, collar end less rounded; cross section rounded or indented. Hatching line inconspicuous. Micropyles in a row on the opercular third, not raised, each located on the junction of carinae between follicular cell impressions (FCIs). Chorion with marked ornamentation of penta- and hexagonal FCIs. Collar round, rim slightly flanged, bears one row of FCIs. Anchor large and flat.
Distribution and biology: Isoperla vevcianensis has a narrow endemic distribution, restricted to the Jablanica Mts, on the border zone of Albania and North Macedonia ( Ikonomov 1980, Murányi 2011). The species is mostly associated with headwater brooks in the alpine and subalpine grasslands, at elevations above 1100 meters ( Figs. 25–27 View FIGURES 25–27 ). The adults emerge in June and July. The adult and larval specimens from our study were found together with Leuctra metsovonica Aubert, 1956 , Perla cf. pallida and another Isoperla species belonging to the I. tripartita species complex (sensu Murányi et al. 2016).
Species relationships: Isoperla vevcianensis was assigned to the Isoperla silesica group sensu Murányi (2011) on the basis of the male penial armatures. Monophyly of the two Central European-Alpine species, I. silesica Illies, 1952 and I. zwicki Tierno de Figueroa & Fochetti, 2001 with the Central Balkanian I. vevcianensis and I. breviptera Ikonomov, 1980 should be confirmed by details of the penial lobes and cox-1 sequences, but these are known only for I. vevcianensis . The mature larva is distinctive among congeners by its large head and short pronotum, similar to the Alpine I. claudiae Graf & Konar, 2014 (in: Graf et al. 2014) and the Crimean I. prokopovi Zhiltzova & Zwick, 2012 . An undescribed species from Montenegro, closely related to I. breviptera , has similar large head and short pronotum (W. Graf, pers. comm.). In the collection of the MM, we have larvae similar to the Montenegrian species, and probably belong to the same undescribed species from Kosovo. The wide trapezoid shaped lacinia is also unique among European species, but this character is rather variable among Nearctic species ( Hilsenhoff & Billmyer 1973, Stewart & Stark 2002, Sandberg 2011). The blunt paraproct is unique among the known larvae of the genus ( Zwick 2004), though this character is not documented for the East Palaearctic and Nearctic species ( Stewart & Stark 2002, Teslenko & Zhiltzova 2006, and Sandberg 2011). For comparison, head, pronotum and paraprocts of sympatric larvae of the unidentified member of the I. tripartita species complex are depicted on Figs. 2 View FIGURES 1–4 , 6 View FIGURES 5–8 , and 8. Larvae of the I. tripartita species complex display characters typical for the West Palaearctic Isoperla . The egg of I. vevcianensis proved to be the usual type of West Palaearctic species, similar to those of e.g. I. tripartita tripartita as depicted in Murányi (2011).
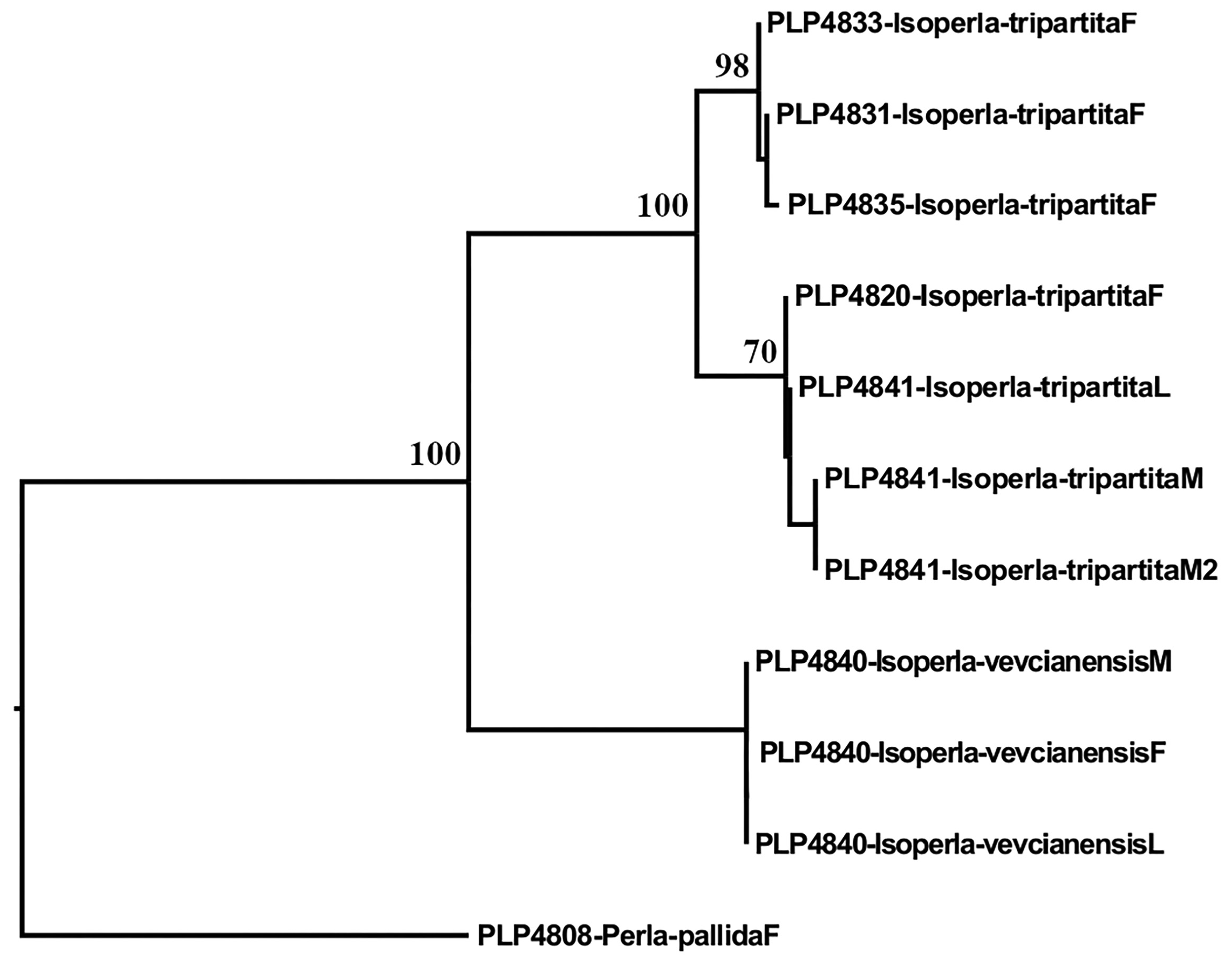
Genetics. Haplotypes from Isoperla vevcianensis formed a cluster of identical sequences for the coxI gene ( Fig. 24 View FIGURE 24 ) with zero intraspecific genetic distance. Molecular analyses agreed with the morphological association between female, male and larvae of I. vevcianensis based on sequence similarities among the specimens. Interspecific genetic distances between I. vevcianensis and the members of the I. tripartita species complex exhibited a 3% divergence, suggesting different species.






| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
| MM |
University of Montpellier |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Isoperla vevcianensis Ikonomov, 1980
| Murányi, Dávid, Kovács, Tibor, Gamboa, Maribet & Watanabe, Kozo 2021 |