
Thermonectus tremouIllesI
|
publication ID |
https://doi.org/ 10.1080/00222933.2016.1145274 |
|
publication LSID |
lsid:zoobank.org:pub:B3603E04-ADE4-4772-A12F-38EB77AC5951 |
|
DOI |
https://doi.org/10.5281/zenodo.5670176 |
|
persistent identifier |
https://treatment.plazi.org/id/5D459B65-FFA7-FFBF-059F-AC39240257C8 |
|
treatment provided by |
Plazi |
|
scientific name |
Thermonectus tremouIllesI |
| status |
sp. nov. |
Thermonectus tremouIllesI View in CoL sp. nov.
( Figures 1 – 4 View Figure 1 View Figure 2 View Figure 3 View Figure 4 )
Type locality
Argentina, Entre Ríos Province, El Palmar NP.
Type material (labels)
Holotype male ( MACN): ‘ Argentina, Entre Ríos Prov., El Palmar NP, 26 – II – 2002, Michat & Torres col. ’ [white, rectangular, printed label], ‘ HOLOTYPE Thermonectus tremouillesi Michat & Torres ’ [red, rectangular, printed label]. Paratypes (28 exs.): same data as holotype [white, rectangular, printed label], ‘ PARATYPE Thermonectus tremouillesi Michat & Torres ’ [red, rectangular, printed label] (13 males, five females, MACN; one male, one female NHML; one male, one female MNHN); same data as holotype, except ‘ 26 – II – 2004 ’ [white, rectangular, printed label], ‘ PARATYPE Thermonectus tremouillesi Michat & Torres ’ [red, rectangular, printed label] (five males, one female, MACN).
Additional material examined
These are damaged or parts of specimens that we prefer not to include in the type series: Argentina, Entre Ríos Province, same data as holotype (six males, three females, MCMC); Misiones Province, Iguazú NP, 12/ 19 August 2000 (one male, MCMC); Misiones Province, Iguazú NP, San Martín Island, 15 February 2002 (1 female, MCMC).
Diagnosis
Medium-sized species; colour pattern on head, pronotum and elytra as in Figure 1 View Figure 1 B, D, E; sexual sculpture of female elytra well visible, covering basal fourth to basal half ( Figure 1 View Figure 1 D); male protibia narrow, dorsal margin slightly prominent ( Figure 2 View Figure 2 D); tergite IX of male with four posterior projections ( Figure 2 View Figure 2 F); median lobe of aedeagus bifid apically ( Figure 2 View Figure 2 E); parameres with a terminal spur ( Figure 2 View Figure 2 E).
Description
Habitus ( Figure 1 View Figure 1 B, C). Oval, greatest width at posterior third, moderately narrowed anteriorly, regularly curved on margins, dorsal surface markedly convex; females somewhat larger than males; male: TL = 11.8 – 12.6 mm (mean = 12.1 mm); MW = 6.7 – 7.4 mm (mean = 7.2 mm) (N = 18) (holotype: TL = 12.4 mm; MW = 7.3 mm); female: TL = 12.2 – 13.2 mm (mean = 12.5 mm); MW = 7.0 – 7.7 mm (mean = 7.3 mm) (N = 6).
Colour. Dorsal surface ( Figure 1 View Figure 1 B, D, E): yellowish to orangish background with black markings as follows: head with postocular band all along posterior margin, projected forward on internal ocular margin to approximately half of eye length, and isolated V-shaped macula in middle of interocular space ( Figure 1 View Figure 1 E), sometimes coalescent with ocular markings ( Figure 1 View Figure 1 B) [holotype: V-shaped macula isolated]; pronotum with transverse band on anterior margin, its width slightly greater than head width, continued posterolaterally in short projections ( Figure 1 View Figure 1 B) that sometimes appear as isolated maculae ( Figure 1 View Figure 1 E) [holotype: anterior band and posterolateral projections connected], and transverse band on posterior margin, a bit narrower and about twice thicker than anterior band, with anterior border indented medially and laterals irregularly rounded and continued to pronotal edge as a fine marginal band; elytra irrorate, irroration absent or reduced on humeral angles, external and internal margins, and two submarginal areas, one just posterior to mid-length, the other subapical; irroration interrupted by an irregular transverse black fascia on distal fourth in some specimens ( Figure 1 View Figure 1 D), in other specimens only an irregular submarginal macula is evident on each elytron ( Figure 1 View Figure 1 B) [holotype: transverse fascia well developed]; internal margin of elytra with a fine black band from scutellum to apex. Ventral surface ( Figure 1 View Figure 1 C): ferrugineous to piceus black except antennae, maxillary and labial palpi, pro- and mesolegs, ventral surface of pronotum, and epipleura yellowish, and base of prosternal process, metatrochantera and distal portion of metafemora orangish.
Sculpture and punctation. Smooth, shiny; microreticulation minute, of regular meshes; head, pronotum and to somewhat lesser degree elytra with very fine, closely distributed punctures combined with larger, sparsely distributed punctures ( Figure 1 View Figure 1 D, SEM detail); each elytron with four longitudinal rows of widely spaced setiferous punctures, one contiguous to elytral commissure, the other three on disc; ventral side with fine, irregularly distributed punctation.
Structure. Head deeply set into pronotum, ratio head length/maximum head width about 0.4; pronotum deeply notched to hold head, ratio length of pronotum at midpoint/greatest width of pronotum about 1/3, ratio width on anterior margin of pronotum/width on posterior margin of pronotum about 0.6; prosternal process moderately widened on distal third, rounded apically ( Figure 2 View Figure 2 C); posterior apex of scutellum rounded; elytra widely rounded on distal third ( Figure 1 View Figure 1 B, D).
Male. Protibia narrow, dorsal margin slightly prominent ( Figure 2 View Figure 2 D); tarsal palette bearing 18 (occasionally 19) sucker-like setae, three of the basal ones a bit larger (among these, one about 20% larger than the other two) and two of the distal ones a bit smaller than the others ( Figure 2 View Figure 2 B); tergite IX with four posterior projections ( Figure 2 View Figure 2 F); median lobe of aedeagus bifid apically ( Figure 2 View Figure 2 E); parameres symmetrical, with a minute, partly concealed apical spur ( Figure 2 View Figure 2 E, detail).
Female. Protibia not distinctly modified, dorsal margin not prominent; elytra bearing well-visible, closely spaced longitudinal sculpture on basal fourth to basal half ( Figure 1 View Figure 1 D).
Variation
The V-shaped macula of head may be more or less developed; in extreme cases it reaches the ocular markings ( Figure 1 View Figure 1 B, E). The pronotal central maculae show different degree of development, from small and isolated ( Figure 1 View Figure 1 E) to somewhat larger and widely connected with the anterior transverse band of the pronotum. The transverse black fascia on distal fourth of the elytra varies greatly in extent; in most specimens it is well developed ( Figure 1 View Figure 1 D), but in some specimens it is reduced and appears as an irregular submarginal macula on each elytron ( Figure 1 View Figure 1 B). The female sexual sculpture varies in extent, covering basal fourth ( Figure 1 View Figure 1 D) to basal half of elytra.
Larvae
Diagnosis. Instar I ( Figures 3 View Figure 3 A – I, 4A – E): head length: 1.40 mm; nasale rounded, markedly projected forward, bearing about 80 – 100 lamellae clypeales on anterior margin ( Figure 3 View Figure 3 A); ratio length of maxillary palpus/length of labial palpus: 0.88 – 0.89; stipes with a row of 15 – 19 additional spine-like setae on dorsal surface ( Figure 3 View Figure 3 E); prementum with three additional spine-like setae on each side of anterodorsal surface ( Figure 3 View Figure 3 H); ratio length of metathoracic leg/head width: 2.59; pro-, meso- and metafemur with one, 1 – 2 and 2 – 3 additional anterodistal setae respectively ( Figure 4 View Figure 4 A); additional measurements and ratios that characterise the body shape are shown in Table 1 View Table 1 .
Instar II: head length: 1.88 mm; nasale rounded, markedly projected forward, bearing about 110 – 130 lamellae clypeales on anterior margin; parietal with 11 temporal and 8 – 9 ventral spine-like setae on each side; antennomere 1 with 5 – 7 secondary hair-like setae; mandible with a row of 30 – 32 secondary hair-like setae on basoexternal margin; ratio length of maxillary palpus/length of labial palpus: 0.88; stipes with a row of 28 dorsal secondary spine-like setae and 16 – 18 external and 35 – 40 dorsoexternal secondary hairlike setae; prementum with 9 – 10 secondary spine-like setae on each side of anterodorsal surface, bearing dorsal spinulae on basal half and on margins; ratio length of metathoracic leg/head width: 2.58; posterior surface of coxa with one secondary pore; additional measurements and ratios that characterise the body shape are shown in Table 1 View Table 1 ; detailed secondary leg setation is given in Table 2.
Instar III ( Figure 4 View Figure 4 F – G): head length: 2.70 – 2.83 mm; nasale rounded, markedly projected forward, bearing about 190 – 210 lamellae clypeales on anterior margin ( Figure 4 View Figure 4 F); anterolateral lobes of frontoclypeus not projecting beyond nasale ( Figure 4 View Figure 4 F); parietal with 11 – 13 temporal and 9 – 15 ventral spine-like setae on each side; antennomere 1 with 8 – 11 secondary hair-like setae; mandible with a row of 28 – 38 secondary hair-like setae on basoexternal margin, and several minute secondary setae on external margin; ratio length of maxillary palpus/length of labial palpus: 0.82 – 0.87; stipes with a row of 25 – 35 dorsal secondary spinelike setae, 17 – 20 external and 32 – 50 dorsoexternal secondary hair-like setae, and few secondary, minute, ventroexternal setae; prementum with 10 – 14 secondary spine-like setae on each side of anterodorsal surface, bearing dorsal spinulae on basal half and on margins ( Figure 4 View Figure 4 G); ratio length of metathoracic leg/head width: 2.68 – 2.82; posterior surface of coxa with 1 – 3 secondary pores; additional measurements and ratios that characterise the body shape are shown in Table 1 View Table 1 ; detailed secondary leg setation is given in Table 2.
Colour. Based on ethanol preserved specimens; therefore, colour pattern might be altered. Instar I: Background colour of cephalic capsule testaceous, stemmatal areas with more or less extended light brown macula; head appendages testaceous, distal half of mandible light brown; thoracic tergites light brown with diffuse testaceous maculae on laterals; legs testaceous to light brown; abdominal sclerites light brown; membranous parts creamy white; urogomphus brown with a narrow ring-like testaceous band at mid length.
Instar II. As for instar I except: light brown stemmatal macula restricted to area surrounding anterolateral stemma; lateral margin of parietal with diffuse light brown area from posterior (smallest) stemma to occipital suture; urogomphus light brown, lacking ringlike testaceous band.
Instar III ( Figure 4 View Figure 4 F). As for instar II except: somewhat darker in general; background colour of cephalic capsule light brown; light brown area on lateral margin of parietal more or less developed (sometimes absent); one small lighter macula contiguous to largest stemma; legs light brown.
Distribution
Argentina, Entre Ríos and Misiones Provinces.
Habitat
Thermonectus tremouillesi apparently exploits a particular habitat. It was found only in lentic water bodies in the forest, shaded with large- and medium-sized trees, with muddy bottom and dark cold water with abundant organic matter. In Iguazú NP (15 February 2002), nine larvae (one of them reared to the adult stage) were collected from a relatively small pond under a bridge, almost completely devoid of aquatic vegetation, in association with adults of Tropisternus Solier, 1834 , Copelatus Erichson, 1832 and Helochares Mulsant, 1844 , and larvae of Tropisternus and Hydaticus palliatus Aubé, 1838 . In El Palmar NP (26 February 2002), several adults and one larva were taken from a small pond 3 m long, 2 m wide and 20 cm in depth, completely devoid of aquatic vegetation, in association with adults of Tropisternus, Enochrus Thomson, 1859 , Copelatus and Rhantus calidus (Fabricius, 1792) . In El Palmar NP (26 February 2004), several adults were taken from a large pond with similar characteristics to the previous sites except for the presence of floating vegetation ( Figure 5 View Figure 5 ), placed contiguously to the small pond sampled in 2002, which by that time was dry.
Etymology
Dedicated to Prof. Edgardo R. Trémouilles, in recognition of his valuable contributions to the knowledge of the dytiscid fauna of South America. The specific epithet is a noun in the genitive case.
Discussion
The identity of many insect species described a long time ago is based on succinct, undetailed and unillustrated original descriptions. Although, by the time they appeared, these descriptions served their purpose, the substantial increase in known diversity during the last few centuries has undoubtedly rendered many old descriptions insufficiently documented from a comparative point of view. This is particularly problematic in the cases in which the holotypes are lost, where species identity might be severely compromised. In the case documented in this paper, after gathering convincing information that the holotype of T. laporti is lost, we considered two possibilities of how to proceed: to consider that our specimens belong to T. laporti and possibly to designate a neotype, or to describe them as a new species. After several discussions with colleagues, the first option appeared more problematic because if we described our material as T. laporti and the lost holotype eventually appeared, there could be some confusion. Also, some presumed differences in dorsal colour pattern (see below) might argue against the recognition of our specimens as T. laporti . On the other hand, if we described our material as a new species, and if the holotype of T. laporti appeared, two scenarios are possible: (1) both holotypes are identical implying a new synonymy, which is no major problem; and (2) both holotypes are different species which means there is no conflict. A careful consideration of these arguments underpinned our decision to describe our material as a new species.
Body size, dorsal colour pattern, and sexual and genital characters have proven to be the most useful in separating species within Thermonectus ( McWilliams 1968; Trémouilles 1989). The distinct dorsal colour pattern of T. tremouillesi differentiates it from most species of the genus. Only four species ( T. alfredi , T. simulator Sharp, 1882 , T. depictus Sharp, 1882 and T. laporti ) exhibit rather similar colouration, mostly on the head and pronotum. Thermonectus alfredi and T. simulator are two closely related species ( Trémouilles 1989), undoubtedly close to T. tremouillesi in aedeagal structure, although visibly larger and bearing completely (or sometimes almost completely in T. alfredi ) irrorate elytra ( Figure 1 View Figure 1 A). The elytra in T. tremouillesi are mostly irrorate but exhibit premarginal yellowish to orangish areas and a more or less extended transverse black fascia ( Figure 1 View Figure 1 B, D) which is never present in the other species. The shape of the male tergite IX of T. tremouillesi , bearing four posterior projections ( Figure 2 View Figure 2 F), clearly differs from that of T. alfredi ( Figure 2 View Figure 2 G) but is rather similar to that of T. simulator . The female sexual sculpture, however, is well marked and closely spaced in T. tremouillesi ( Figure 1 View Figure 1 D), whereas it is very subtle and more sparsely distributed in T. simulator . Also, the male protibia is narrow, with the dorsal margin slightly prominent in T. tremouillesi ( Figure 2 View Figure 2 D), whereas in T. simulator and T. alfredi it is broader and the dorsal margin is more prominent, giving the tibia a rather triangular appearance ( Trémouilles 1989). On the other hand, in T. depictus , the anterior pronotal transverse band has pointed ends, and the posterolateral projections (or isolated central maculae) are absent, whereas the tergite IX of the male bears only two posterior projections ( Trémouilles 1989). As already mentioned, the separation of T. tremouillesi and T. laporti is more difficult based on available information. Both species have similar size and general colouration, although the following differences can be marked after comparison of our specimens with the original description of T. laporti ( Aubé 1838) : (1) the projection forward (following the internal ocular margins to approximately half of the eye length) of the postocular black band, which is present in T. tremouillesi , is not mentioned for T. laporti ; this is a rather conspicuous feature, and its omission from the description of the latter species may indicate an absence; (2) the posterior transverse black band of the pronotum is 2.5 times broader than long in T. laporti , whereas it is more than four times broader than long in T. tremouillesi ; also, the laterals of this band are continued to the pronotal edge as a fine marginal band in T. tremouillesi , which is not mentioned for T. laporti ; (3) the elytra of T. laporti are described as being black and covered with yellowish rounded maculae; in T. tremouillesi the background colour of the elytra is yellowish to orangish and covered with black irroration. This last character, however, should be taken with care as it is just a point of view which colour (black or yellow) to choose as the background. Thermonectus tremouillesi keys to couplet 6 in Tremouilles ’ (1989) key on South American Thermonectus . This couplet is formed by T. simulator and T. alfredi , both species differing from T. tremouillesi in the characters mentioned above.
Despite great morphological similarity in larval morphology among Thermonectus species, the larvae of T. tremouillesi look more similar to those of T. alfredi ( Michat 2013) than to those of T. succinctus ( Michat and Torres 2005) . The nasale is rounded and somewhat more projected forward in all instars of T. tremouillesi and T. alfredi ( Figures 3 View Figure 3 A, 4F). In T. succinctus , the nasale is more flattened and less prominent. The first instars of T. tremouillesi and T. alfredi have a larger number of additional spine-like setae on the dorsal surface of the stipes (15 – 19 and 17 – 21 respectively, Figure 3 View Figure 3 E) and bear additional anterodistal setae on the femur ( Figure 4 View Figure 4 A). Thermonectus succinctus , on the other hand, has fewer (6 – 12) additional setae on the stipes and lacks additional anterodistal setae on the femur. The second and third instars of T. tremouillesi can be distinguished from those of T. alfredi and T. succinctus in the larger number of secondary spine-like setae on the anterodorsal surface of the prementum. Also, the number of secondary hair-like setae on the basoexternal margin of the mandible is larger in T. tremouillesi and T. alfredi than in T. succinctus . With respect to the third instar, in T. tremouillesi and T. alfredi , the basal one-half to two-thirds of the dorsal surface of the prementum is covered with spinulae ( Figure 4 View Figure 4 G), whereas in T. succinctus , the spinulae are present only on the basal third. The dorsal colour pattern of the cephalic capsule in T. tremouillesi and T. succinctus lacks light maculae on each side of the frontoclypeus and on the parietals (close to the joint of frontal and coronal sutures), thus differing from T. alfredi . The absence of dark maculae on the frontoclypeus of the mature larva of T. tremouillesi ( Figure 4 View Figure 4 F) also distinguishes it from the mainly North American species T. basillaris ( Harris, 1829) and T. nigrofasciatus ornaticollis ( Aubé 1838) , which bear, in different degrees, frontoclypeal dark maculae ( Wilson 1923; Carroll and Barman 2004).

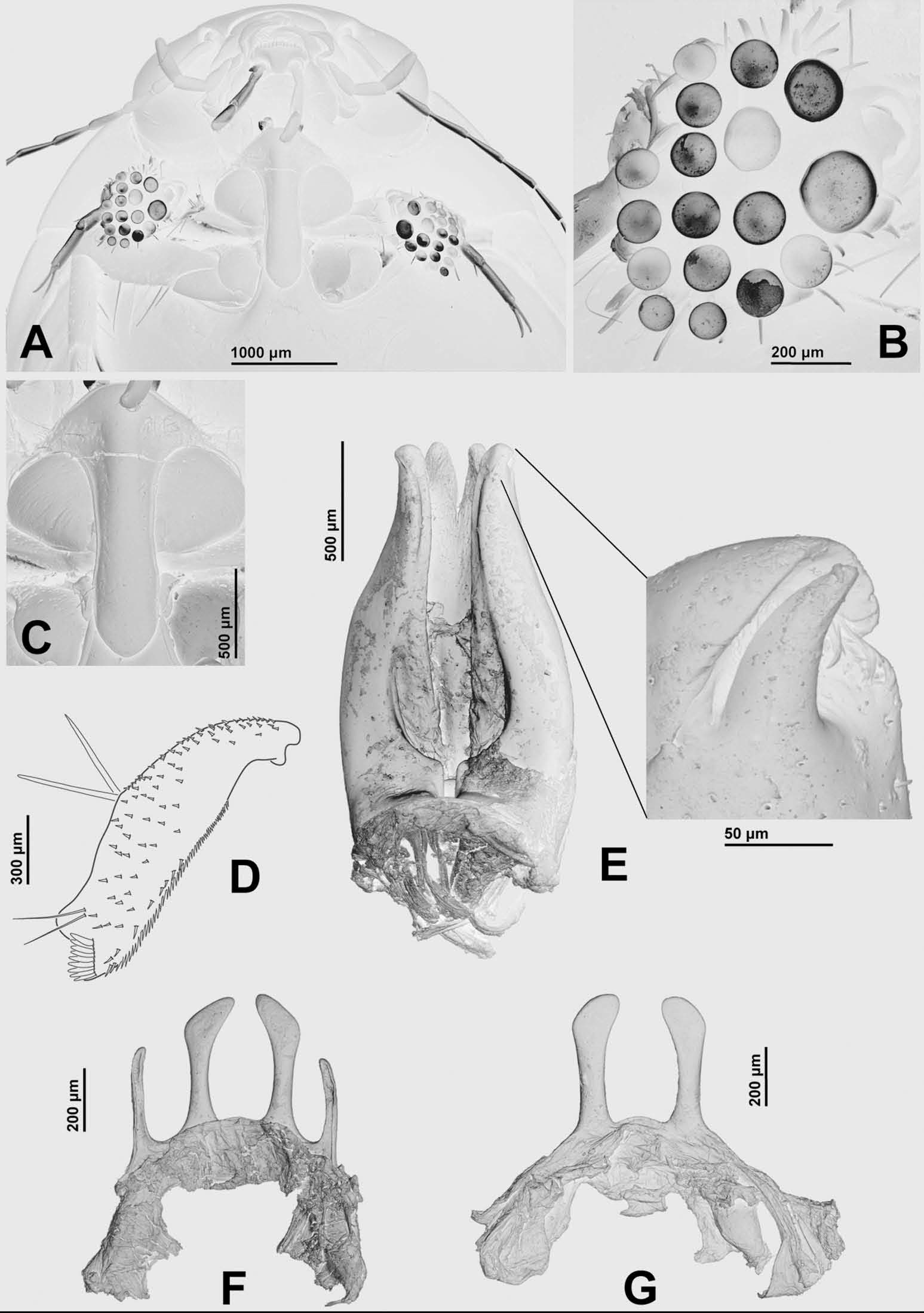
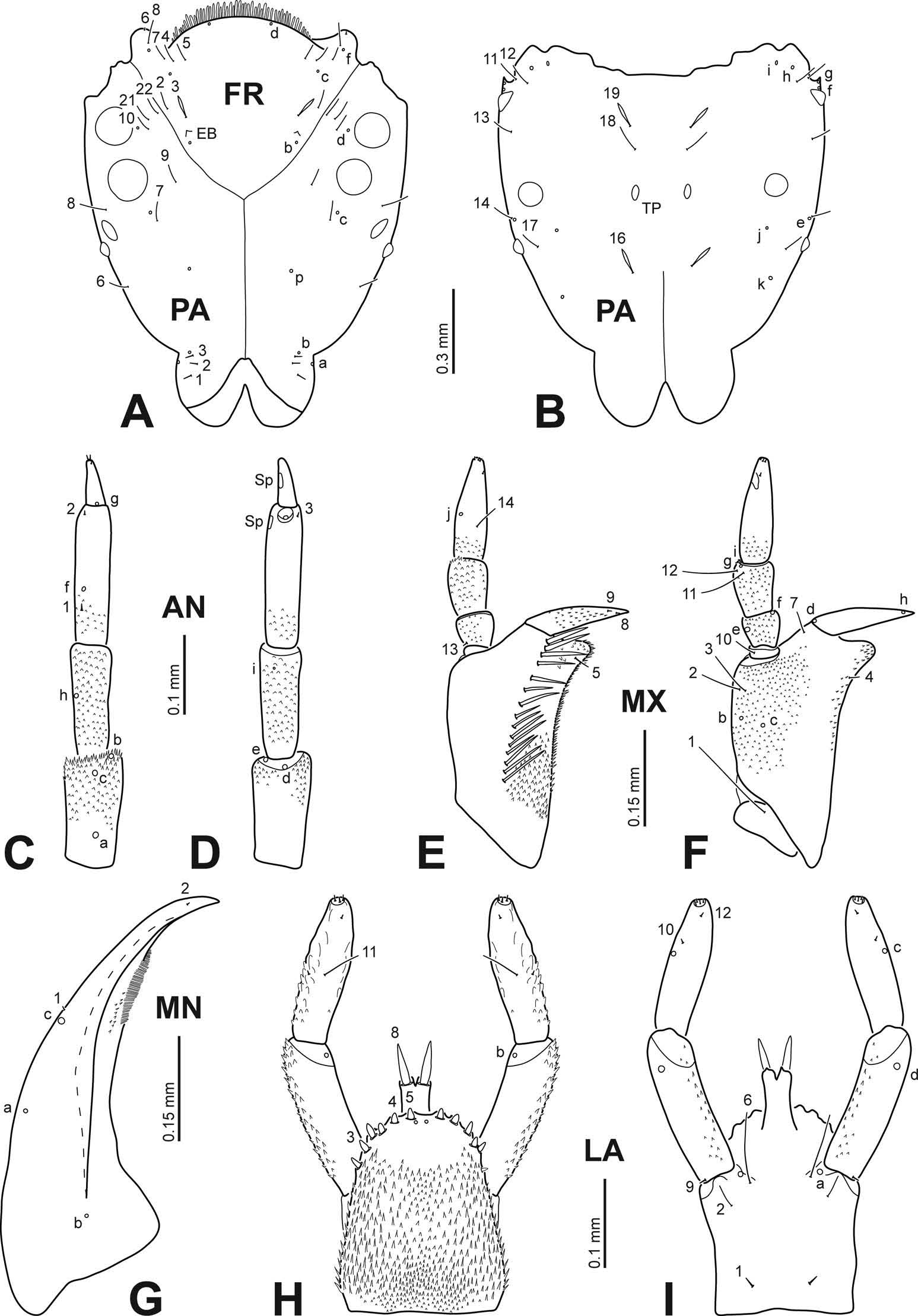
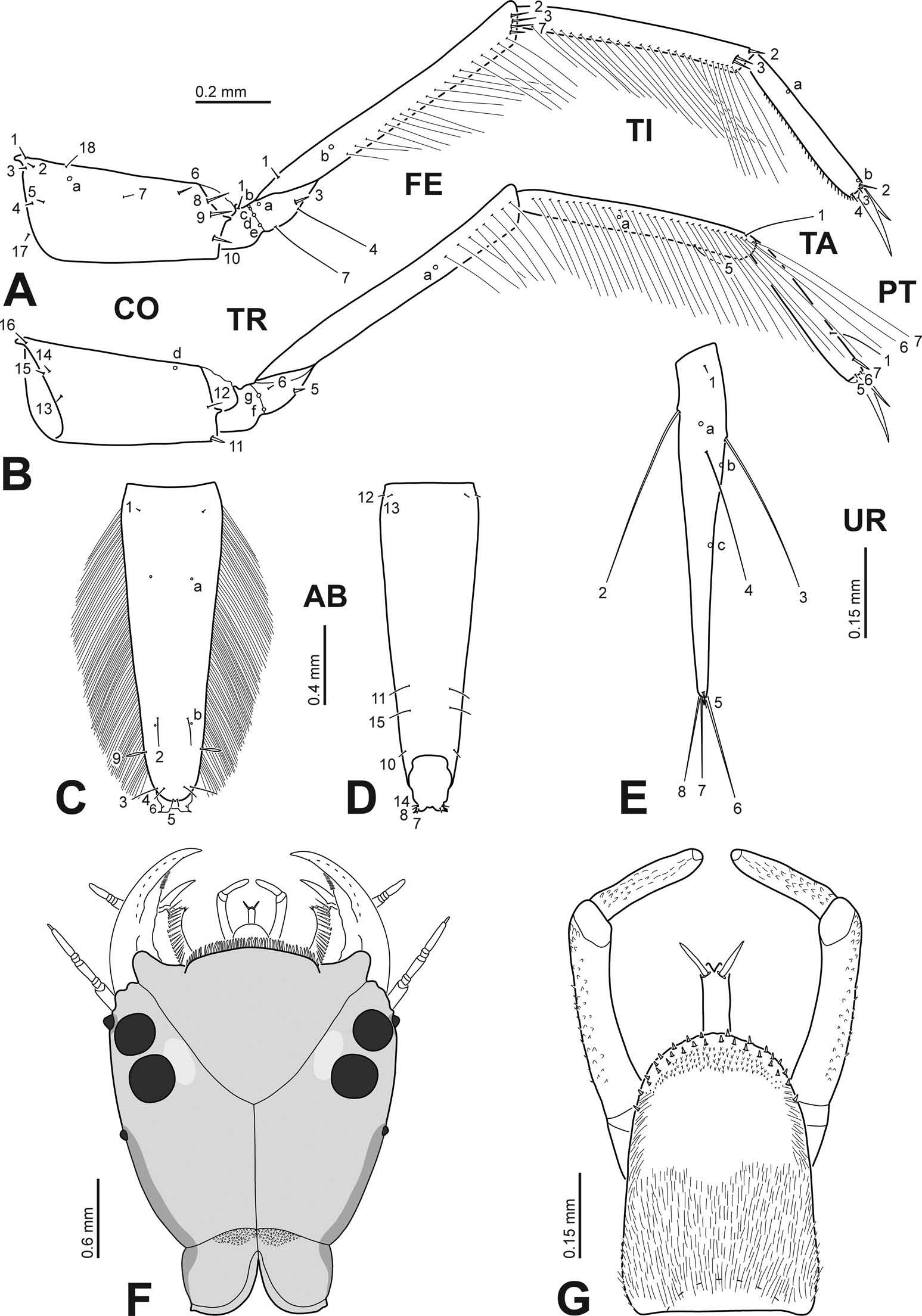

Table 1. Measurements and ratios for the larvae of Thermonectus tremouillesi.
| Measure | Instar I (n = 2) | Instar II (n = 1) | Instar III (n = 5) |
|---|---|---|---|
| TL (mm) | 8.00–8.30 | 13.10 | 15.20–23.60 |
| MW (mm) | 0.80–1.20 | 1.60 | 2.10–2.90 |
| HL (mm) | 1.40 | 1.88 | 2.70–2.83 |
| HW (mm) | 1.15 | 1.56 | 2.16–2.21 |
| FRL (mm) | 0.64–0.65 | 0.84 | 1.16–1.22 |
| OCW (mm) | 0.43 | 0.56 | 0.92–0.99 |
| HL/HW | 1.22 | 1.20 | 1.23–1.28 |
| HW/OCW | 2.71 | 2.78 | 2.22–2.38 |
| COL/HL | 0.54 | 0.55 | 0.56–0.57 |
| FRL/HL | 0.46 | 0.45 | 0.43–0.44 |
| A/HW | 0.50 | 0.44 | 0.37–0.42 |
| A1/A3 | 0.79 | 0.79 | 0.71–0.86 |
| A2/A3 | 0.79–0.82 | 0.88 | 0.81–0.93 |
| A4/A3 | 0.33–0.37 | 0.25 | 0.16–0.20 |
| MNL/MNW | 2.74–2.89 | 3.04 | 3.13–3.30 |
| MNL/HL | 0.46 | 0.47 | 0.44–0.46 |
| PPF/MP1 | 0.36–0.40 | 0.22 | 0.20–0.25 |
| A/MP | 1.77–1.81 | 1.79 | 1.58–1.76 |
| MP1/MP2 | 0.56–0.58 | 0.75 | 0.76–0.91 |
| MP3/MP2 | 1.84–1.89 | 1.50 | 1.29–1.45 |
| GA/MP1 | 2.91–3.20 | 2.61 | 2.06–2.48 |
| MP/LP | 0.88–0.89 | 0.88 | 0.82–0.87 |
| LP2/LP1 | 0.94–0.95 | 0.82 | 0.64–0.72 |
| L3 (mm) | 2.86–2.98 | 4.03 | 5.81–6.25 |
| L3/L1 | 1.00–1.03 | 1.07 | 1.11–1.12 |
| L3/L2 | 0.95–0.97 | 0.99 | 1.01–1.02 |
| L3/HW | 2.59 | 2.58 | 2.68–2.82 |
| L3 (CO/FE) | 0.74–0.81 | 0.76 | 0.81–0.86 |
| L3 (TI/FE) | 0.76–0.77 | 0.72 | 0.65–0.70 |
| L3 (TA/FE) | 0.58–0.59 | 0.52 | 0.45–0.48 |
| L3 (CL/TA) | 0.36–0.37 | 0.25 | 0.23–0.24 |
| LAS (mm) | 1.55–1.63 | 1.93 | 2.68–3.00 |
| LAS/HW | 1.41 | 1.23 | 1.22–1.33 |
| U (mm) | 0.60 | 0.75 | 1.05–1.34 |
| U/LAS | 0.37–0.39 | 0.39 | 0.39–0.45 |
| U/HW | 0.52 | 0.48 | 0.49–0.59 |
A: antenna; A1–A4: antennomeres 1–4; CL: longest claw; CO: coxa; COL: coronal line length; FE:
femur; FRL: frontoclypeus length; GA: galea; HL: head length; HW: head width; LAS: length of last
abdominal segment; LP: labial palpus; LP1–LP2: labial palpomeres 1–2; L1–L3: legs 1–3; MNL:
mandible length; MNW: mandible width; MP: maxillary palpus; MP1–MP3: maxillary palpomeres
1–3; MW: maximum body width; OCW: occipital foramen width; PPF: palpifer; TA: tarsus; TI: tibia; TL:
total body length (excluding urogomphi); U: urogomphus length.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.