
Araiocypris batodes, Conway & Kottelat, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.4509070 |
|
persistent identifier |
https://treatment.plazi.org/id/5D388569-FFBF-A850-FEB1-FE65BBB3CBE8 |
|
treatment provided by |
Carolina |
|
scientific name |
Araiocypris batodes |
| status |
sp. nov. |
Araiocypris batodes View in CoL , new species
( Figs. 1, 2 View Fig )
Gen. et spec. ind.: Kottelat, 2001: 45, Fig. 68
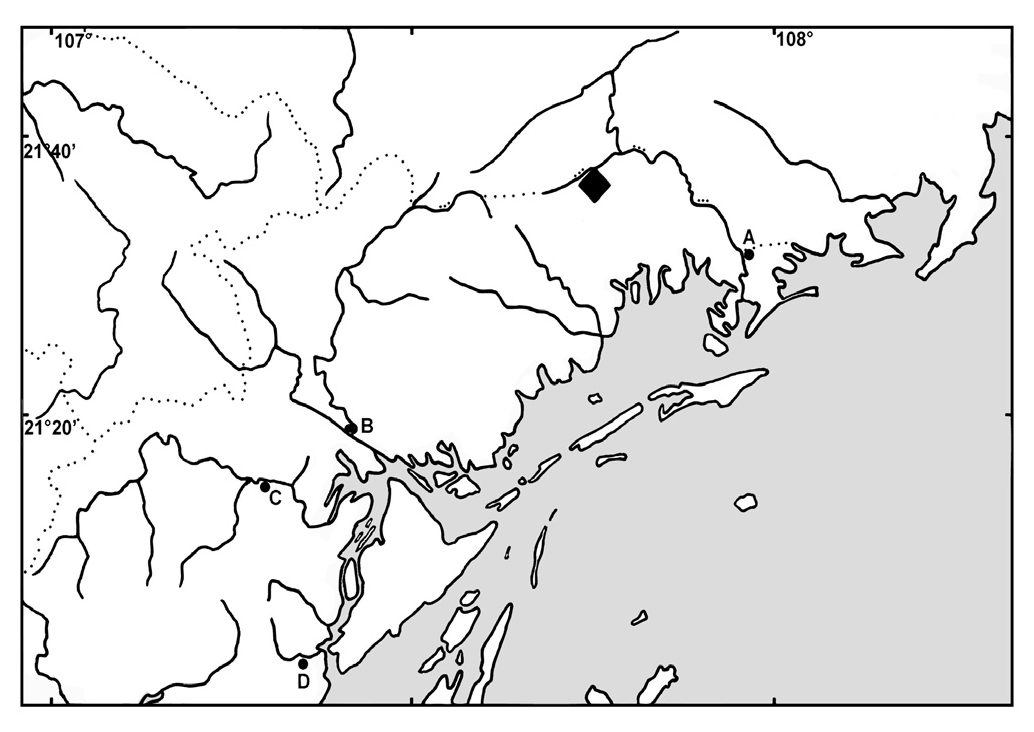
Holotype. – ZRC 50942, male, 33.2 mm SL; Vietnam: Quang Ninh Province: Hai Ninh District: forest creek near Khay Rau village, 10 km on road from Bac Phong Sinh to Mong Cai ; 21 ° 33'44"N 107 ° 43'55"E; M. Kottelat et al., 2 Oct.1998. GoogleMaps
Paratypes. – CMK 14899, female, 30.5 mm SL; same data as holotype GoogleMaps .
Diagnosis. – See generic diagnosis.
Description. – Small cyprinid fish, maximum size 33.2 mm SL. General body shape as in Figs. 1 and 2 View Fig . Morphometric characters of the two known specimens (holotype listed first) as follows: head length 20.5, 21.6% SL, predorsal length 62.0, 61.9% SL, preanal length 60.2, 60.3% SL, prepelvic length 45.1, 45.9 % SL, body depth at dorsal-fin origin 14.7, 16.7% SL, caudal peduncle length 15.0, 15.9% SL, caudal peduncle depth 7.2, 6.8% SL, length of anal-fin base 22.6, 24.2% SL, length of dorsal-fin base 8.4, 9.8% SL, eye diameter 33.8, 33.3% HL, snout length 17.6, 19.7% HL. Dorsal fin with ii.5.i, ii.6.i rays, anal with iii.15.i, iii.16. Principal caudal-rays 9+8, dorsal procurrent rays 5, 6, ventral procurrent rays 6. Pelvicfin rays i.4.i, pectoral-fin rays i.10.i. Eye large, orbit diameter 33.3, 33.8% HL, mouth superior, lower jaw projecting further anteriorly than snout tip. Anterior and posterior nostril openings widely separate, anterior opening at tip of a short tube. Body elongate with short dorsal fin, origin level or slighty posterior to anal-fin origin. Caudal fin forked. Pelvicfin origin anterior to dorsal-fin origin. Pectoral fin large, falciform. Muscles associated with pectoral-fin rays greatly enlarged, particularly abductor superficialis and arrector ventralis. Small dorsoventrally elongated window in hypaxial musculature at lateral side of anterior swimbladder chamber, extending along posterior edge of cleithrum.
Two distinct rows of tubercles present along lower jaw of male, outer row consisting of two or three large conical tubercles. Inner row consisting of three or four smaller rounded tubercles. Numerous minute tubercles speckled evenly across chest region, dorsal surface of head and along lower jaw; single row of minute tubercles also forming a complete ring around eye of male. Female with minute tubercles speckled across chest region, dorsal surface of head and along lower jaw only. Strip of minute tubercles present along dorsal surface of unbranched portion of branched pectoral-fin rays 2–4 in male only.
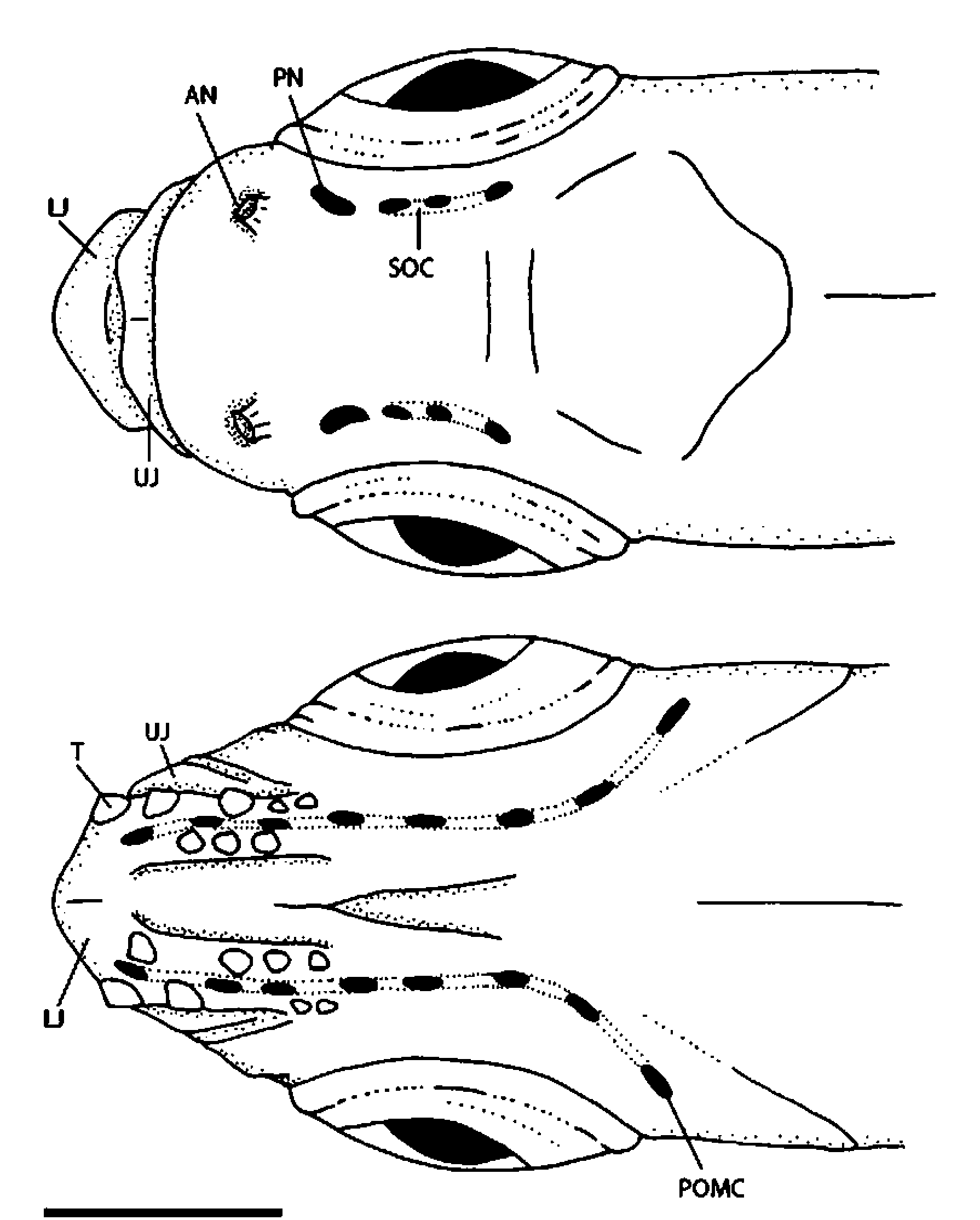
Cephalic sensory system reduced to preoperculo-mandibular canal and supraorbital canals. Sensory canal openings large, 8 in preoperculo-mandibular canal (3 on lower jaw, 5 on preopercular), 3 in supraorbital canal. Lateral line absent. Scales small, transparent, irregularly arranged, 49–50 along midlateral row, 1 / 2 /3/ 1 / 2 transverse scale rows on caudal peduncle.
Colouration in life. – Body pale yellow with a faint brown or reddish brown midlateral stripe. Black spot at caudal-fin base. Caudal fin yellow. The first impression in the field was of a colour pattern identical to that of Tanichthys albonubes .
Colouration in alcohol. – Body background colour whitish, with a large black spot on lower half of caudal-fin base. Narrow depigmented stripe along entire length of body, dorsal to horizontal septum, originating posterior to opercle and continuing to caudal peduncle, bordered ventrally along entire length by a broad swath of melanophores, densest anteriorly, thinning out towards posterior. Scale pockets dorsal to depigmented stripe lightly edged with dark melanophores, forming a reticulate pattern. Narrow but distinct axial streak along horizontal septum, ventral to depigmented stripe, starting just anterior of anal-fin origin and continuing to caudal spot. Narrow but distinct row of small melanophores along entire length of dorsal midline, from neurocranium posteriorly to caudal fin. Dorsal area of occipital region heavily speckled with large melanophores, continuous with dorsal pigment row. Dorsal surface of snout between nostrils lightly speckled with small melanophores. Also a light scattering of large melanophores on lachrymal and opercle. Dense speckling of melanophores on lower jaw. Dark row of large melanophores running along base of anal fin, continuing along ventral edge of caudal peduncle. Thin streaks of small melanophores along principal caudal fin rays and along branched rays of dorsal, anal, pectoral and pelvic fins.
Distribution. – Araiocypris batodes is presently known only from the type locality, a forest creek, apparently in the Ka Longo drainage ( Fig. 3 View Fig ); the exact drainage could not be determined on site or from available maps, and local information was contradictory. Ka Longo marks the border between Vietnam and China and is known as Beilun He in China. The habitat was a small stream in the foothills, and
THE RAFFLES BULLETIN OF ZOOLOGY 2008
Araiocypris batodes was collected in a shallow (less than 40 cm deep) bay with slow water current and sandy bottom, adjacent to riffles. Despite intense efforts to obtain more, only two specimens were obtained.
Etymology. – From the Greek βατοδης´(batodes), thorny, like a blackberry (βατομουρο´). Treated here as a noun in apposition. In allusion to the large conical tubercles on the lower jaw of the male.
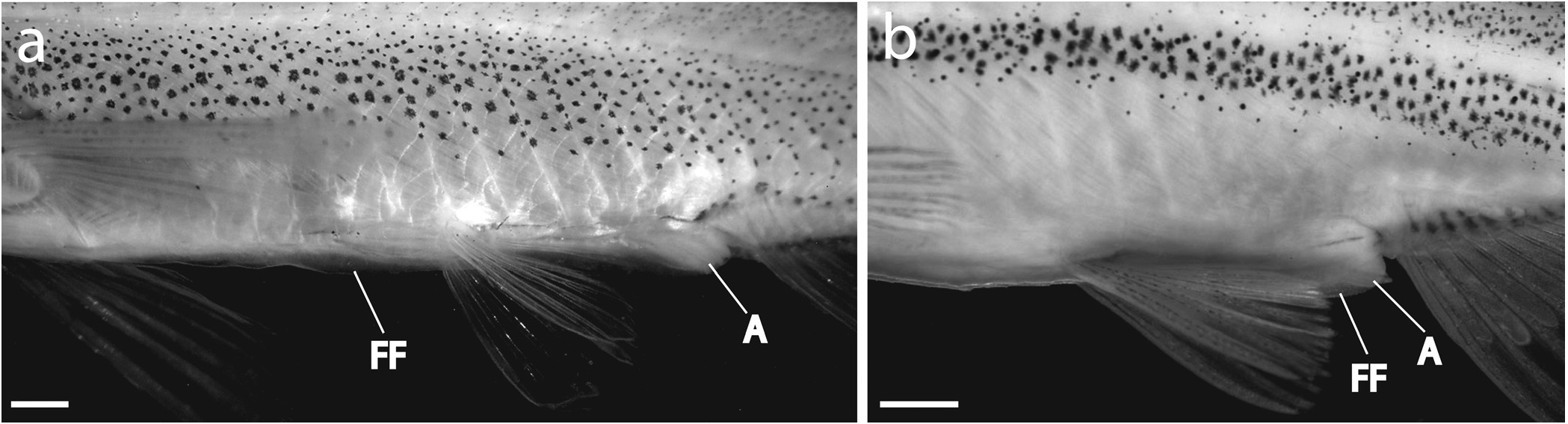
Remarks. – This species cannot be assigned to any of the known genera of Southeast and East Asian cyprinids and it is therefore placed in a new genus. The combination of a soft and flexible pre-anal fin-fold, extending from the pectoral region to the anus, and the tubular anterior nostril appears to be an as yet unique combination of characters.
Several small cyprinid species are known to possess a soft and flexible pre-anal fin-fold, a remnant of the pre-anal larvalfin-fold, including Paedocypris progenetica Kottelat et al. 2006 , Danionella mirifica Britz, 2003 , and Hemigrammocypris lini Weitzman & Chan, 1966 (they referred to the fin-fold as a ventral abdominal keel; p. 288). All these are species of very small adult body size (less than 30 mm SL). In these species, the fold does not reach far in front of the pelvics and therefore differs from the condition in Araiocypris batodes where the soft and flexible fin-fold extends past the pelvics reaching almost to pectoral-fin insertion ( Fig. 5a View Fig ).
exposed which is also referred to as a ‘scaleless keel’ ( Freyhof & Kottelat, 2007; Kottelat & Freyhof, 2007). A similar ‘scaleless keel’ is also reported as present in some species of Metzia , referred to as a postventral keel ( Berg, 1932: 156) or an abdominal keel ( Xie et al., 2003: 403).
These are all different from the condition found in the new species, and clearly all ‘keels’ and ‘fin-folds’ are not strictly homologous. In Paedocypris progenetica the soft and flexible pre-anal and post-anal fin-folds are remnants of the pre-anal and post-anal larval-fin-folds respectively, which are retained in the developmentally truncated adult ( Kottelat et al., 2006). It is plausible to assume that the pre-anal and post-anal folds present in Danionella mirifica are the products of similar developmental truncations. We do not see reasons to believe that the hard and inflexible keel-like structure extending along the ventral midline between the pectoral region and anus in some members of the Palaearctic and Oriental genera of the leuciscine, cultrine and xenocyprine lineages are homologous with the pre-anal fin-fold of Paedocypris progenetica as this keel-like structure develops at a later stage in ontogeny, after the pre-anal larval-fin-fold has been lost. The fold extending along the ventral midline of Araiocypris batodes is similar in proportions to the keel-like structure of many Palaearctic and Oriental genera of the leuciscine, cultrine and xenocyprine lineages but differs in being soft and flexible (vs. hard and A keel-like structure of similar proportions is also found in several Palaearctic and Oriental genera of the leuciscine, cultrine and xenocyprine lineages, including Pseudolaubuca , Toxabramis , Cultrichthys , Parabramis and Hypothalmichthys (see Chen, 1998). However, in these taxa the keel is hard and inflexible (L. Yang, pers. comm.; pers. obs.). A keel (a sharp, compressed belly) is also present in many species of leuciscines, chelines and rasborines but this structure is covered by scales. In a few species of the Western Palaearctic genus Alburnus , some of the scales along the ventral midline in front of the anus are missing leaving a short skin-fold inflexible) and in this regard more akin to the pre-anal finfold present in species of Paedocypris and Danionella . Without access to a development series of Araiocypris batodes it is difficult for us to definitively state whether the soft and flexible fold extending along the ventral surface of Araiocypris batodes is a remnant of the larval-fin-fold or a superficially similar structure that develops later in ontogeny.
The only species within a similar size range to Araiocypris batodes and possessing a soft and flexible pre-anal fin-fold that we are aware of is Hemigrammocypris lini ( Fig. 5b View Fig ). Though both species lack barbels and a lateral line and exhibit a reduced cephalic sensory system ( Fig. 4 View Fig ), there is no reason to hypothesise a close relationship as these features are common to many small fish species and are thus of limited use in resolving the interrelationships of small fish genera ( Conway, 2005; Britz & Moritz, 2007). Furthermore, Araiocypris is easily distinguished from Hemigrammocypris by possessing more branched anal-fin rays (15–16 vs. 9–11), more midlateral scales (49–50 vs. 30–33) and fewer pelvic fin rays (i,4,i vs. i,6–7).
The general features and relative positioning of the nostrils of Araiocypris batodes are unlike those of other small cyprinid species known to us. The anterior nostril is a tube-like opening and is widely separated from the smaller posterior nostril, which is situated posterior to the anterior margin of the orbit. In most other small rasborine taxa that we have examined the anterior nostril is usually rimmed by a short, fleshy wall, which may or may not exhibit a nostril barbel or flap, and the posterior nostril (usually larger than the anterior nostril) is usually situated immediately posterior to the anterior nostril. In certain species ( Tanichthys albonubes , Tanichthys micagemmae , Barboides britzi and Barboides gracilis ) the anterior and posterior nostrils are confluent and form a figure eight shaped opening (Weitzman & Chan, 1966; Freyhof & Herder, 2003; Conway & Moritz, 2006). This later condition is likely due to a developmental truncation in the formation of the skin over the nasal cavity, which usually forms the separation between the anterior and posterior nostrils (R. Britz, pers. comm.).
The presence of minute tubercles on the head and chest of both specimens of A. batodes , the minute tubercles on unbranched pectoral-fin rays 2–4 of the male and the large conical tubercles arranged serially on the lower jaw of the male suggests that the two specimens have reached or are close to attaining adult size. The tubercles on the lower jaw of the male are similar in terms of arrangement (single outer row of large conical tubercles, inner row(s) of smaller conical tubercles) to those of males of some species of Zacco , Parazacco , Candidia and Opsariichthys (e.g. see Kottelat, 2001: Fig. 41; Chen & Chang, 2005: Figs. 17, 28, 41) but the large falciform pectoral-fins and the keeled appearance of the ventral surface are more reminiscent of cultrines and some rasborines (e.g. Chela , Oxygaster ; Howes, 1979, 1980). As only two specimens of Araiocypris batodes are known we could not carry out a survey of its osteology. We therefore restrict ourselves to attracting attention to this new species and decline from making speculations about its possible affinities at this time.
Comparative material. – Barboides britzi, BMNH 2005.6.7.44, holotype, 11.5 mm SL. – BMNH 2005.6.7.45– 57, 13 paratypes, 10.8–11.6 mm SL. – BMNH 2005.6.7.63– 68, 6 paratypes, 11.3–12.9 mm SL; Benin: Lokoli Swamp Forest. Barboides gracilis, BMNH 1970.9.24.2, neotype and holotype of Barbus lorenzi , 13.1 mm SL; BMNH 1970.9.24.3–4, 2 paratypes of Barbus lorenzi , 13.7–14.0 mm SL; Benin: Iguidi River at Gbokoutou.– BMNH 2005.6.7.1– 41, 41 specimens, 7.3–11.9 mm SL; Benin: Iguidi River at Langbay. – AMNH 97625, 15 specimens; Cameroon: tributary of Lobe River near Lobe Falls, Bomaa. Danionella mirifica, USNM 372848, 36 paratypes, 10.9–14.1 mm SL; Myanmar: Kachin Division: hill stream, 8 miles from Kamaing on road to Tanai. Paedocypris micromegethes: BMNH 2004.11.16.1–40, 40 specimens, 9.7–11.6 mm SL; Borneo: Sarawak: Batu Kawa-Matang, Taman Koperkasa, ca. 10 km from Kuching. Paedocypris progenetica: CMK 11260, 28 paratypes, 7.8–10.3 mm SL; Sumatra: Jambi Province: swamp near Pematang Lumut, 40 km before Kuala Tungkal on road to Jambi (95 km) and Simpangtuan (36 km). Sundadanio axelrodi: BMNH 1982.3.29.50–54, 5 specimens, 15.3–17.5 mm SL; Indonesia: Borneo, Kapuas River at Kapuas. – BMNH 2005.11.10.1–8, 8 specimens, 15.7–20.3 mm SL; locality unknown. – BMNH 2005.11.10.9–16, 8 specimens, 17.7–19.7 mm SL; locality unknown. – CMK 8424, 44 specimens, 8.6–19.7 mm SL; Malaysia: Sarawak: Blackwater stream in forest at km 7 on road from Kuching to Batu Kawa. – CMK 9633, 39 specimen, 10.6–19.0 mm
THE RAFFLES BULLETIN OF ZOOLOGY 2008
SL; Indonesia: Sumatra: Banka , 5.5 km North of Payung on road to Pangkalpinang. – CMK 10876, 29 specimens, 10.2– 18.1 mm SL; Malaysia : Sarawak : Sungai Nibung , about 1 km North of Durin ferry on Sungai Rajang on road from Sri Aman to Sibu. – CMK 10978, 5 specimens, 14.7–17.5 mm SL; Malaysia : Sarawak : Sungai Tebu , km 8 on raod from Daro to Matu. – CMK 16731, 23 specimens, 12.1–17.3 mm SL; Indonesia : Borneo : Kalimantan Selatan, area of Tamiyang Layang, Barito River Basin. Tanichthys albonubes, BMNH 1935.4.18.30, 1 specimen; China : Canton : Kwangtung, South China . – BMNH 1936.12 .1.3, 1 specimen; China : White Cloud Mountain , Canton. Hemigrammocypris lini, USNM 359591, 1 specimen, 30 mm SL; locality unknown. – USNM 197762 View Materials , 1 specimen, 35 mm SL; China : Hong Kong .

| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.