
Gnathia nasuta Nunomura, 1992
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3737.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:CE23AED0-EBA7-422D-AB3C-47DC2D3FB4D |
|
DOI |
https://doi.org/10.5281/zenodo.6160799 |
|
persistent identifier |
https://treatment.plazi.org/id/412987E8-FFB6-3311-FF3C-FE95BFB42E97 |
|
treatment provided by |
Plazi |
|
scientific name |
Gnathia nasuta Nunomura, 1992 |
| status |
|
Gnathia nasuta Nunomura, 1992 View in CoL
(Japanese name: Hanadaka-umi-kuwagata) ( Figs 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Gnathia nasuta Nunomura, 1992: 69 –71, fig. 6.—Nunomura, 1995: 209, fig. 21-196G.—Saito et al. 2000: 17.
Caecognathia nasuta .—Cohen & Poore 1994: 288.—Smit & Davies 2004: 296.—Nunomura & Shimomura 2012: 585, 586, fig. 99.
Gnathia hirayamai Nunomura, 1992: 65 –67, fig. 4.—Cohen & Poore 1994: 289.—Nunomura, 1995: 209, fig. 21- 196 F.—Saito et al. 2000: 17.—Tanaka 2004: 58.—Nunomura, 2004: 354.—Nunomura & Shimomura 2013a: 142, 145.—Nunomura & Shimomura 2013b: 292, fig. 132. [New synonymy]
Gnathia nagasakiensis Nunomura, 2012: 62 , 63, fig. 3.—Nunomura & Shimomura 2013a: 142, 144.—Nunomura & Shimomura 2013b: 293, fig. 134. [New synonymy]
Gnathia shijikiensis Nunomura, 2012a: 64 , 65, fig. 4.—Nunomura & Shimomura 2013a: 142, 144.—Nunomura & Shimomura 2013b: 294, fig. 135. [New synonymy]
Gnathia brevicephala Nunomura, 2012a: 65 –67, fig. 5.—Nunomura & Shimomura 2013a: 142, 144.—Nunomura & Shimomura 2013b: 293, fig. 133. [New synonymy]
Material examined of type series. Male adult, holotype of Caecognathia nasuta (Nunomura, 1992) , TOYA Cr- 11172, 3.3 mm, from 8.5 m depth, off Tomioka, Reihoku-cho, Kumamoto Pref., Kyushu, western Japan (32°20– 22′N, 130°01–03′E), 22 May 1980, coll. Taiji Kikuchi. Adult male, holotype of Gnathia hirayamai Nunomura, 1992 , TOYA Cr-11167, 3.3 mm, from same locality and date of G. na s u t a holotype.
Male adult, holotype of Gnathia nagasakiensis Nunomura, 2012 a, TOYA Cr-23312 (in original description but actually Cr-23352, 5.0 mm in original description but actually 3.9 mm), from 23 m depth of Shijiki Bay, Nagasaki Pref., western Japan (33°11′–13′N, 129°24′–21′E), 11 May 1984, coll. Mikio Azuma. Male adult, holotype of Gnathia shijikiensis Nunomura, 2012 a, TOYA Cr-23353, 5.6 mm in original description but actually 4.4 mm, from 35 m depth of Shijiki Bay, 11 May 1984, coll. Mikio Azuma. Male adult, holotype of Gnathia brevicephala Nunomura, 1992 , TOYA Cr-23356, 5.6 mm in original description but actually 4.3 mm, from 25 m depth of Shijiki Bay, 2 May 1984, coll. Mikio Azuma. All materials from Shijiki Bay were collected by Nagasaki University Sledge (NUS) net or Smith-McIntyre grab.
New material. Two male adults, TOYA Cr-23570 and 23571, 3.1 and 3.1 mm, from the same locality and date of C. nasuta holotype. One male adult, KMNH IvR-500,708, 2.4 mm, from 180 m depth off east Amami-oshima Island, southern Japan (28°15.00′N, 129°35.00′E), sledge net, 10 Nov. 1994, coll. Takeo Yamauchi, TR /V Toyoshio-maru. Six male adults, KMNH IvR-500,709, 2.2–3.2 mm, from 126 m depth, sandy and muddy sediment, off west Mi-shima Island, Sea of Japan (34°36.163′–138′N, 130°32.750′–894′E), sledge net, 18 Nov. 1999, coll. Michitaka Shimomura, TR /V Toyoshio-maru. One male adult, KMNH IvR-500,710, 2.6 mm, from 163–170 m depth, off north Tokashiki-jima Island, Kerama Islands, southern Japan (26°03.67′N, 127°21.14′E), beam trawl, 26 May 2006, coll. Michitaka Shimomura, TR /V Toyoshio-maru. Two male adults, KMNH IvR- 500,711, 2.3 and 2.3 mm, from 71–75 m depth, off south Okinawa-jima Island, southern Japan (26°04.50′N, 127°32.22′E), beam trawl, 26 May 2006, coll. Michitaka Shimomura, TR /V Toyoshio-maru. One male adult, KMNH IvR-500,712, 3.2 mm, from 403–412 m depth, off southeast Amami-oshima Island, southern Japan (27°58.02′N, 129°24.03′E), inner net of beam trawl, 22 May 2009, coll Susumu Otsuka, TR /V Toyoshio-maru. Two male adults, KMNH IvR-500,713, 2.5 and 3.0 mm, from 130 m depth, muddy sediment with foraminiferal shells, off west Goto Islands, western Japan (33°04.8′N, 128°20.2′E), benthos net, 25 Jul. 2010, coll. Keiich Kakui, R/V Soyo-maru. Two male adults, KMNH IvR-500,714, 2.7 and 3.4 mm, from 142 m depth, muddy sediment with foraminiferal shells, off west Goto Islands, western Japan (33°16.1′N, 128°20.2′E), benthos net, 25 Jul. 2010, coll. Keiich Kakui, R/V Soyo-maru. 17 male adults, KMNH IvR-500,715, 2.4–3.8 mm, from 213 m depth, sandy and muddy sediment, off east Amakusa Town, Kumamoto Pref., western Japan (32°13.4′N, 128°39.1′E), dredge, 26 Jul. 2010, coll. Keiichi Kakui, R/V Soyo-maru. Nine male adults, KMNH IvR-500,716, 1.9–3.0 mm, from 205 m depth of Tosa Bay, Shikoku, southern Japan (33°14′45.249′′N, 133°37′54.048′′E), sledge net with 1 mm cod end, 24 Oct. 2012, coll. Tomomi Saito. Six and 1 male adults, KMNH IvR-500,717 and 500,718, 2.9–3.6 mm, from 63.1–75.9 m and 24.8–29.2 m depth, muddy sediments, off Tateyama, Boso Peninsula, central Japan (34°59.47′N, 139°46.98′E and 34°59.66′N, 139°48.50′E), dredge, 16 Apr. 2013, coll. Yuzo Ota, TR /V Hosaka-maru.
Distribution ( Fig. 1 View FIGURE 1 , Table 1). From southwestern to central Japan (8.5–412 m depth).
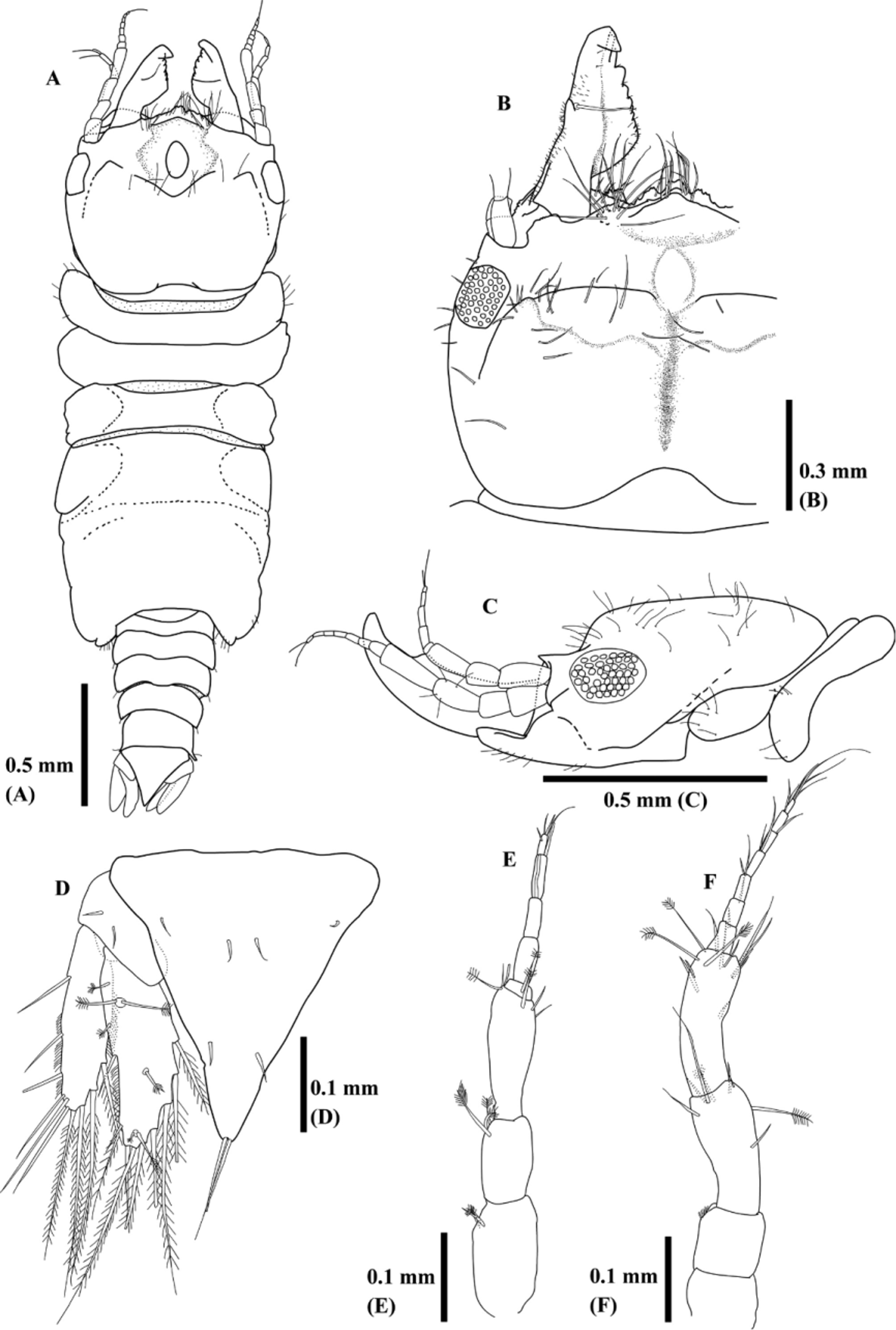
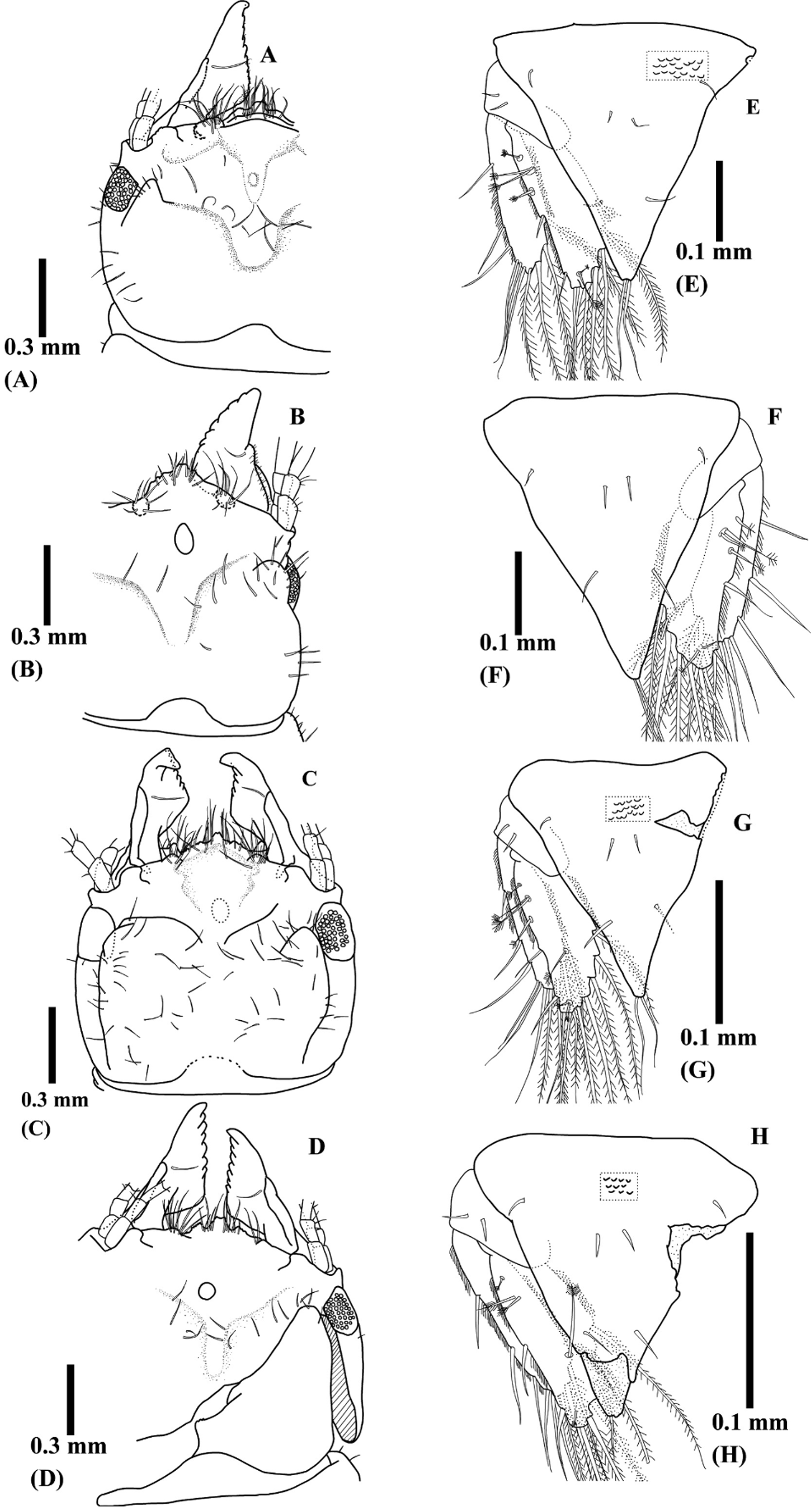
Redescription ( Figs 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 ). Body 1.9–3.8 mm (mean ± SD; 2.8 ± 0.4 mm, n = 51, new materials) without tubercles. Color of fixed specimens white or light brown; digestive organs brown due to congealed host blood. Cephalosome ( Figs 5 View FIGURE 5 A–C, 7A–D) almost square with posterior margin slightly concave, sparsely covered with setae. Anterior part of cephalosome acutely lowered in thickness at near eyes in lateral view. Frontal border convex with many setae. Mediofrontal process and 2 frontolateral processes visible on central frontal border and apex smooth or slightly dentate. Mediofrontal process extends 2 frontolateral processes. Superior frontolateral processes with several setae. Tips of ventral frontal border often visible under superior frontolateral processes. Protrusions apparently visible inner sides of basis of antennae. Low and pentagonal rising often visible near frontal border. Dorsal sulcus narrow or U-shaped. Oval-shaped translucent region visible anteromedianly on dorsal sulcus. Eyes composed with 44 ocelli. Paraocular ornamentation prominent as one protrusion. Supraocular lobe not prominent with smooth apex.
Pereonite I ( Figs 5 View FIGURE 5 A–C, 7A–D) short, not fused or medially fused with cephalosome, reaching lateral margin of cephalosome. Widths and lengths of pereonites II, III, and IV almost same. Pereonites II and III anteriorly concave. Pereonite IV without constriction and anterolateral lobes slightly visible. Areae laterales slightly visible on pereonites V. Pereonite VII short and narrow, overlapping pleonite I.
Pleonite I–IV ( Fig. 5 View FIGURE 5 A) almost equal length and narrower posteriorly, with 1 seta on lateral margin of each segment. Pleonite V longer than each segment of Pleonite I–IV. Epimera not prominent.
Pleotelson ( Figs. 5 View FIGURE 5 D, 7E–H) almost triangular with acute apex and often covered with scales. Three pair of seta on dorsal surface and 1 pair of seta on distal apex.
Mandible ( Figs. 5 View FIGURE 5 A–C, 7A–D) nipper-like shape, seems as triangular in dorsal view, from half to two-third length of cephalosome. Apex often bends backward. Dentate blade occupies over half-length of mandible. One mandibular seta presents near armed carina on mid-dorsal surface. Basal neck and erisma prominent.
Antennule ( Fig. 5 View FIGURE 5 E) composed with 3 peduncle and 5 flagellar articles. Two, 3, and 2 feather-like bristles on distal margins of peduncle article I, II, and III, respectively. One feather-like bristle on flagellar article I. One aesthetasc on flagellar article III and IV, and V, respectively. Flagellar article V terminating in 3 setae.
Antenna ( Fig. 5 View FIGURE 5 F) composed with 4 peduncle and 7 flagellar articles. One and 3 feather-like bristles on peduncle III and IV, respectively. 0–2 setae on distal margins of flagellar article I–VI. Article VII terminates in 4 setae.
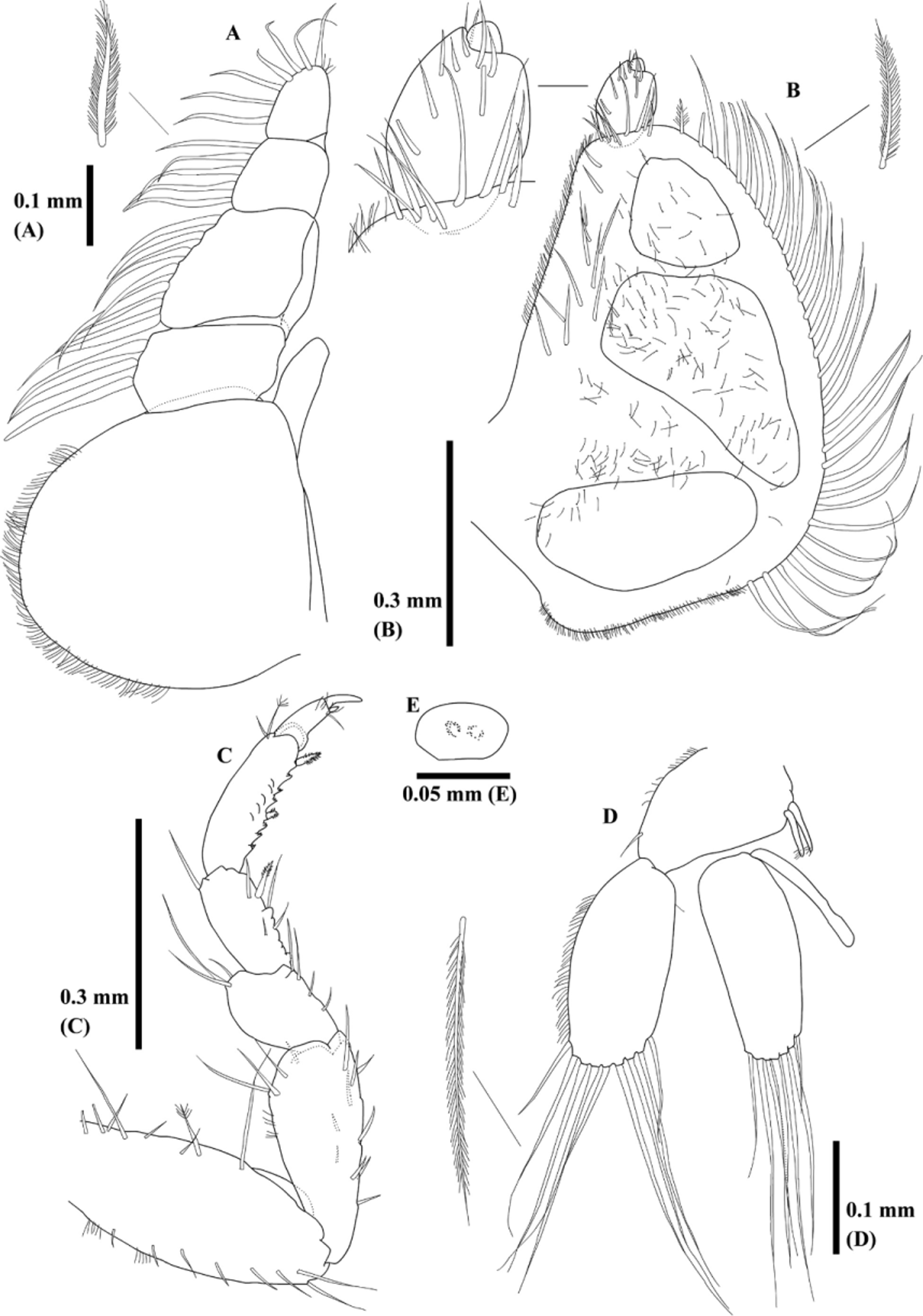
Maxilliped ( Fig. 6 View FIGURE 6 A) with endite reaching palp article II. Three, 7, 5, and 7 plumose setae on external margins of palp articles I–IV, respectively. Four setae on internal margin of palp article IV.
Pylopod ( Fig. 6 View FIGURE 6 B) 3-articled. Article I semicircular with posterior corner; 3 areolae visible, 32 plumose setae on internal margin, 12 setae on external margin, 9 setae on distal margin, and fine setae cover ventral surface. Article II elliptical with 10 setae. Article III minute.
Pereopod II ( Fig. 6 View FIGURE 6 C) sparsely covered with setae. Basis oblong with 1 feather-like bristle. Ischium shorter than basis, becoming larger distally. Merus about half-length of ischium. Carpus slightly larger than merus with 1 denticulate compound spine on inner margin. Propodus rectangular, bearing 2 denticulate compound spines on inner-mid and inner-distal margins. Dactylus terminating in unguis.
Pleopod II ( Fig. 6 View FIGURE 6 D) with 1 seta on outer distal corner and coupling hook on inner margin. Both rami oval and equal in length. Appendix masculina of pleopod II extending half-length of endopod. Eight or 9 plumose setae on exopods. Seven or 8 plumose setae on endopods.
Uropod ( Figs. 5 View FIGURE 5 D, 7E–H,) fringed with fine setae laterally. Both rami subequal in length. Exopod not extend beyond apex of pleotelson and endopod almost reach apex. Exopod with 6 simple and 4 plumose setae and endopod with 3 simple and 6 plumose setae, and several feather-like bristles.
Penes ( Fig. 6 View FIGURE 6 E) composed with 2 contiguous papillae.
Remarks. Caecognathia nasuta and Gnathia hirayamai were described by Nunomura 1992, and G. nagasakiensis , G. shijikiensis , and G. brevicephala were described by Nunomura, 2012a. A redescription of their five holotypes revealed that they shared the following characteristics: nipper-shaped mandibles; dentate blade occupying more than half the length of the mandible (apparently triangular in shape in the dorsal view); acutely lowered anterior part of the cephalosome; convex frontal border with several setae; mediofrontal process extending into two frontolateral processes; superior frontolateral processes with several setae; prominent paraocular ornamentation as one protrusion, and acute apex of the pleotelson.
The following are the differences between the drawings by Nunomura (1992) and the present study. Caecognathia hirayamai : (1) “a conspicuous incisor” was mentioned in the original description but it was inconspicuous in the present study; (2) the frontal border was almost straight without processes but there was actually a convex frontal border with several setae and three frontal processes; (3) the number of flagellar articles of the antenna was mentioned as nine but it was actually seven; (4) article III of the pylopod was not drawn. Caecognathia nasuta : (1) a “big pentagonal frontal border” was drawn but there was actually a convex frontal border. Furthermore, Nunomura (1992) did not describe pereonite I and the endite of the maxilliped, although they were actually present.
The following are the differences between the drawings by Nunomura (2012a) and the present study. Gnathia nagasakiensis : (1) the frontal process and setae were not clearly shown; (2) “small rings” were described, which were actually absent. Gnathia shijikiensis : (1) pereonite I was not drawn. Gnathia brevicephala : (1) the number of flagellar articles of the antennule and antenna were mentioned as five and six, but there were actually five and seven, respectively; (2) article I was described as oval-shaped but it was actually semicircular. Furthermore, Nunomura (2012a) did not describe the endite of the maxilliped, although it was actually present. Prominent paraocular ornamentation was not observed in the original descriptions of these five species. Considering the similarities between E. kikuchii and its synonyms, the present study determined that G. hirayamai , G. nagasakiensis , G. shijikiensis , and G. brevicephala were junior synonyms of Caecognathia nasuta .
The present drawing of the G. nagasakiensis holotype ( Fig. 7 View FIGURE 7 B) appears to be different from other specimens with respect to the mandible shapes and eyes, the basal neck and armed carina are not distinct, the tip of the mandible is not curved inward, and the eyes are presented laterally. This is because the mandibles are closed and the basal neck is hidden behind the frontal border, while the armed carina is visible laterally. The cephalosome of this specimen was deformed during the previous dissection and the location of the right eye moved laterally (the left eye was located dorsally). This specimen was preserved with other male materials (non type) and their morphologies were similar. Thus, G. nagasakiensis is considered to be a junior synonym of C. nasuta .
The genus of the present species was considered to be Caecognathia by Cohen & Poore (1994) but it belongs to the genus Gnathia because of the following features: the frontal border is not excavated and has frontal processes; paraocular ornamentation; a dorsal sulcus is present on the cephalosome.
Gnathia nasuta was closely related to C. nipponensis (Monod, 1926) in terms of the following characteristics: nipper-shaped mandible, prominent paraocular ornamentation as one protrusion, acute apex of the pleotelson, and appendix masculina reaching half the length of the endopod of pleopod II. Caecognathia nipponensis was not regarded as Gnathia by Cohen & Poore (1994). However, the original description may have failed to reveal the frontal processes on the frontal border (Fig. 149 in Monod 1926). More detailed observations of the C. nipponensis holotype and morphological comparisons will be necessary in future.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |