
Listriodon splendens VON MEYER, 1846
|
publication ID |
https://doi.org/ 10.37520/fi.2020.021 |
|
persistent identifier |
https://treatment.plazi.org/id/3C2287B8-FFC8-FFE0-FF6C-86FC8621FB14 |
|
treatment provided by |
Felipe |
|
scientific name |
Listriodon splendens VON MEYER, 1846 |
| status |
|
Listriodon splendens VON MEYER, 1846
L e c t o t y p e. MHNCHF 8, right I1/ in the Natural History Museum, La Chaux-de-Fonds, figured by Stehlin (1900: pl. 5, fig. 22). Lectotype designated by Van der Made (1996: 100, pl. 41, fig. 1a, b, c and 2c – the latter is mislabelled as MHNCHF 8’ left I1/). The specimen has lost part of the root since being figured by Stehlin (see also Bayle 1856).
P a r a l e c t o t y p e s. According to Van der Made (1996): MHNCHF 8’, left I1/; MHNCHF 56b, left c/ 1m; MHNCHF 10, left i/3; MHNCHF 4, left P4/; MHNCHF 9aua, left I1/; MHNCHF 48, right C1/m ( Calydonius tener VON MEYER, 1846 ); MHNCHF 10x, left i/3; MHNCHF 2, right m/3; MHNCHF 4, left P2/; MHNCHF 10, right i/3; MHNCHF 6, left i/1; MHNCHF 6, right i/1; MHNCHF 45, right C1/m (one of the syntypes of Calydonius trux VON MEYER, 1846 ).
Material associated with the lectotype. According to Van der Made (1998), plate 1: MHNC 48 View Materials , left C1/m (contradiction of side with Van der Made 1996) ; MHNC 9 View Materials aua, left I1 /; MHNC, right cuboid; MHNC 6 View Materials , right i/1 ; MHNC 10 View Materials x, left i/3 ; MHNC, left calcaneum; plate 2: MHNC, right astragalus; MHNC, left navicular; MHNC, right magnum.
A note on the subdivision of the s p e c i e s L i s t r i o d o n s p l e n d e n s. Van der Made (1996) proposed a tripartite subdivision of Listriodon splendens with chronological connotations, the lineage being interpreted as increasing in dimensions over geological time from small Listriodon splendens tapirotherium (DE BLAINVILLE, 1847) to medium-sized Listriodon splendens splendens VON MEYER, 1848 , and thence to large Listriodon splendens major ROMAN, 1907 . He also referred to this sequence as evolutionary stages I, II and III. Stage I was considered to be typical of early MN 6, stage II characteristic of the transition MN 6–7, whereas stage III was reported to occur at the transition MN 8–9. Pickford and Morales (2003) reported the presence of a huge specimen from Toril, Spain, that challenged this scheme because Toril is correlated to MN 7, but in general, the sequence appears to be sound.
The naming of the large subspecies of Listriodon splendens is problematic in that some of the material from La Chaux-de-Fonds is large, falling into the range of variation of L. s. major, yet it has a prior name trux VON MEYER, 1846 . The available evidence suggests that trux rather than major is likely to be the valid name for this subspecies. The differences in dimensions of the sample from La Chaux-de-Fonds could reflect sexual bimodality rather than the presence of two species or subspecies, but the available material is too restricted to resolve the issue. In essence, the problem is one of nomenclature. Should L. s. major be called L. s. trux ?
M a t e r i a l f r o m N e b i s u y u. EUNHM PV 3075, skull lacking the premaxillae, and left maxilla fragment containing the canine.
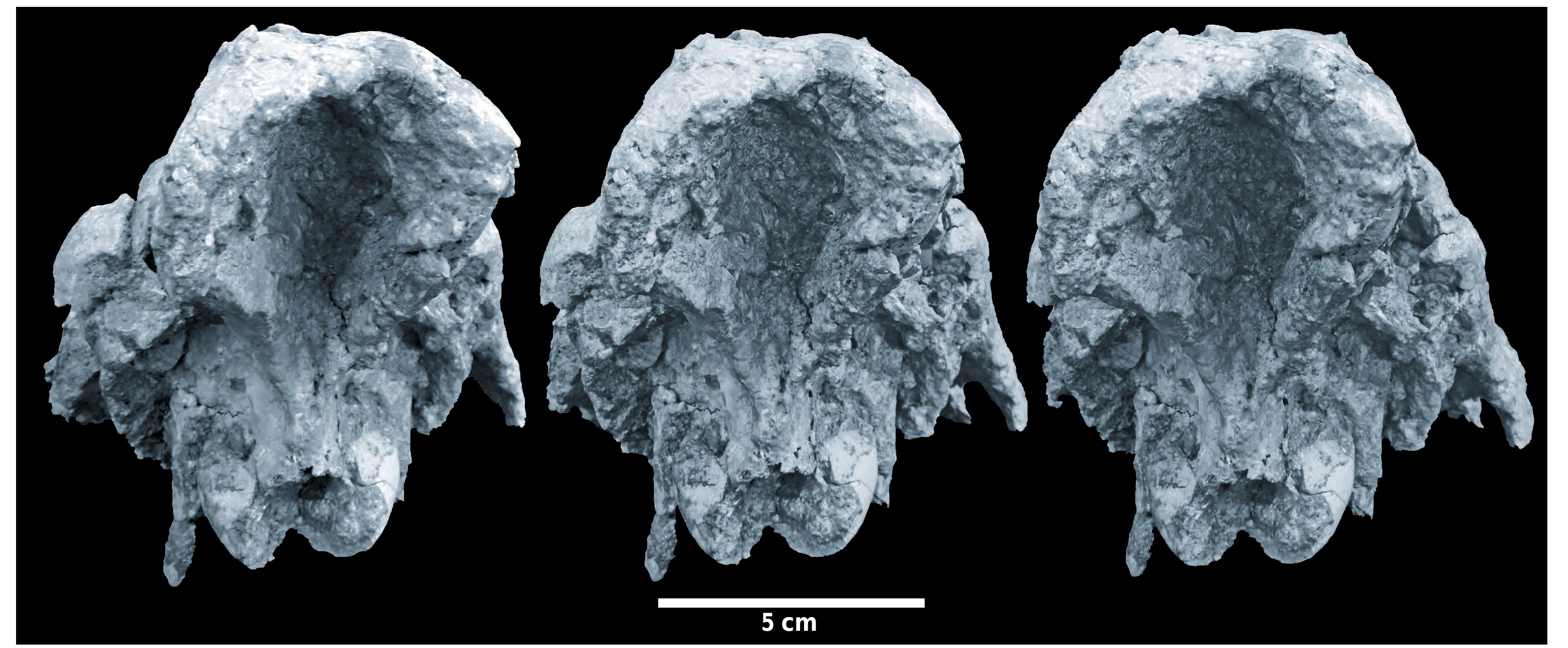
D e s c r i p t i o n. Skull. In dorsal view ( Text-fig. 2 View Text-fig ), the frontal bone dominates the skull, not only by its relatively great breadth, but also by its massiveness, its somewhat rugose surface and the thickness of the bone, extending even onto the post-orbital process. The anterior parts of the frontal bones narrow markedly and descend towards the nasals at a steep angle, and are scored by two deep, parallel, grooves that link distally to the supra-orbital foramina. The supra-orbital foramina are poorly exposed but lie about half way down the steeply sloping anterior surface of the frontal. The frontals are domed upwards between the lachrymals, and are so swollen that they overhang the latter bones to a small extent. Careful scrutiny of the frontal dome reveals the presence of a suture all around the top of the frontal, on top of which is a bony apophysis, the apex of which has broken off. The broken base of the apex is oval and measures 50 mm antero-posteriorly by 31 mm bilaterally. This bony apophysis resembles an ossicone rather than a horn, since its origin appears to have been dermal. The lachrymal foramina are not preserved.
The parietals posterior to the frontals are not swollen and the surface of the bones is smooth. The temporal crests are far apart, but because the skull is slightly crushed, there is an irregular ridge running along the midline, but this should not be confused with a sagittal crest, although it could represent a distal extension of the suprafrontal structure or ossicone. The posterior margin of the neurocranium is incurved rather than rectilinear.
The distal extremities of the zygomatic arches are visible in dorsal view, and they terminate about half way between the posterior margin of the orbit and the nuchal ridge at the rear of the skull.
Finally, in dorsal view, it is possible to make out that the zygomatic arches slope laterally and ventrally to a marked extent, but because the lateral edges of the structures are broken off on both sides it is not possible to determine the diameter of the skull at this point, nor whether the zygoma possessed laterally projecting apophyses. They do, however, have a rugose upper surface and are dorso-ventrally thickened, which is rather unusual in a suid context.
In lateral view ( Text-figs 3 View Text-fig , 4 View Text-fig ), it is once again the frontal bones that dominate the skull. They comprise a solid dome-like structure above the lachrymals, the swollen and somewhat rugose bone surface extending posteriorly onto the post-orbital processes, which are themselves enlarged antero-posteriorly and thicker than is usual in suid skulls. The suture between the frontal and the lachrymal is well exposed on both sides. The anterior parts of the frontals descend steeply towards the nasals before curving anteriorly. The nasals are broken off.
The anterior margin of the orbits is about 13 mm behind the distal extremity of the M3/ and are positioned high in the skull. Distortion makes it difficult to estimate the original dimensions of the orbits.
Posteriorly, the temporal fossae are clear and deeply excavated. The zygomatic arches are large and form a massive base to the orbits. However, their lateral extremities have been broken off, so it is not possible to ascertain their original extent.
The occipital condyles are small and lie only slightly above the level of the plane of the cheek teeth and well beneath the level of the orbits. The paroccipital processes are short and are directed ventro-distally.
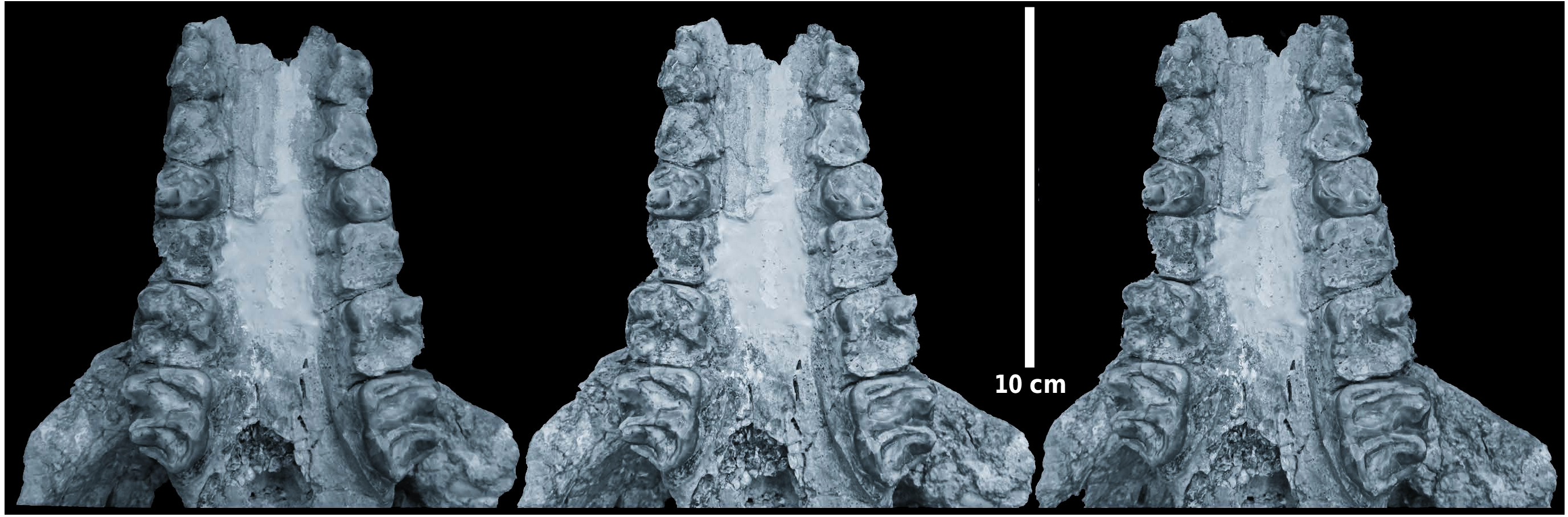
In ventral view ( Text-fig. 5 View Text-fig ), the palate is observed to extend beyond the distal ends of the M3/s by about 22 mm (estimated due to damage). The zygomatic arches more than double the breadth of the skull, but to what extent is difficult to estimate due to the fact that the lateral parts of both zygoma are broken off. The root of the zygomatic process of the maxilla is far back, opposite the front of M3/. The tooth rows diverge gently from fore to aft, but the skull is slightly crushed and the palate has been partly reconstructed so this divergence may be somewhat altered by post-mortem distortion. The occipital condyles appear to be small for such a massive skull, and the paroccipital processes are short and are oriented ventro-distally. Much of the basicranium is preserved, but the state of preservation does not permit accurate description or interpretation.
In supero-anterior view ( Text-fig. 6 View Text-fig ), there are two aspects of the skull which are striking to the eye. The first is the narrowness of the snout, the second is the massiveness of the frontal zone which dominates the area above the orbits. The two grooves linked to the supraorbital foramina are deep and subparallel, coursing along the dorsal part of the frontals. The foramina from which they emerge are poorly preserved, partly due to damage, but partly due to thickening of the frontal bone which has a distinctly rugose surface which contrasts vividly with the smooth bone of the lachrymals beneath them.
In supero-anterior view the base of the ossicone structure is clearly visible, forming an apophysis above the thickened and domed frontal bone and marked by a suture (subtle but clearly visible in glancing light). This ossicone is located above and anterior to the orbits in the anterior part of the frontal dome, the main part of which lies between the orbits. In this view, the thickening of the post-orbital processes is clearly evident, the bones forming a massive margin at the rear of each orbit. This contrasts with the sharp edges of the rear extremities of the zygomatic arches which form the margins of the temporal fossae.
In distal view, the point that leaps to the attention is the deeply concave nuchal area of the neurocranium. The upper margins of the depression are thickened laterally and dorsally, and gradually diminish in thickness as they approach the occipital condyles, where they form a V-shaped junction in the mid-line of the skull.
In posterior view ( Text-fig. 7 View Text-fig ) the paroccipital processes are short and oriented vertically just to the lateral side of the occipital condyles. They extend barely a cm beyond the lowest point of the condyles.
Dentition. The palate retains heavily worn P2/–M3/ on both sides ( Text-fig. 8 View Text-fig ).
The P2/ and P3/ are worn to stubs revealing little about their occlusal cusp morphology. On the buccal sides of both the P2/ and the P3/, there is a low, beaded cingulum, while a depression on the lateral surface of each tooth indicates that there was likely a metacone in these two teeth, now largely eliminated by wear. The occlusal outline of each tooth is triangular with rounded corners. P4/ is heavily worn but shows three cusps, the protocone and paracone forming an anterior loph, and the metacone somewhat isolated in the disto-buccal corner of the crown. There are remnants of mesial and distal cingula. M1/ and M2/ are reduced to stubs with no cuspal morphology discernible. The M3/ in contrast, even though heavily worn shows two clear transverse lophs separated by a transverse valley which is not blocked by a median accessory cusplet. There is a mesial cingulum, especially evident in the mesio-lingual half of the crown, and there is a distal cingulum between the metacone and hypoconule. Wear has advanced to the stage where the protocone and paracone are joined together thereby forming a strong, slightly oblique transverse loph, and the metacone, hypocone and hypoconule are joined together, forming an obliquely oriented distal loph.
The overall aspect of the dentition of the Nebisuyu suid skull is that it is of a highly lophodont listriodont such as Listriodon splendens , but it is not as finely lophodont as Lopholistriodon kidogosana PICKFORD et WILKINSON, 1975 , which is in any case a much smaller animal ( Pickford 1986). Metrically, the teeth of the Nebisuyu fossil plot into the upper part of the range of variation of dentitions of Listriodon splendens from Europe ( Text-fig. 9 View Text-fig ), with the sole exception that the breadth of the P2/ slightly surpasses that of any other known specimen. However, considering its extremely worn status, measurement of the P2/ had to be estimated from the roots which are known to be larger than the crown. Therefore, the seemingly very broad P2/ does not comprise a substantive argument against attributing the specimen to Listriodon splendens .
If one accepts the presence of diverse subspecies of Listriodon splendens , as was the position of Van der Made (1996) then, on the basis of its large cheek teeth with a suggestion that P2/ and P3/ possessed distinct metacones, one would conclude that the Nebisuyu skull should be classified as Listriodon splendens major ROMAN, 1907 , or as Listriodon splendens Evolutionary Stage III of Van der Made (1996). Given, however, that the type specimen of Calydonius trux VON MEYER, 1846 , is a large upper canine from the same locality as the upper central incisor which is the lectotype of Listriodon splendens , the possibility exists that evolutionary stage III should be called either splendens or trux , both names having priority over major. The question is delicate in that the lectotype incisor, even though large within the context of Listriodon , could represent evolutionary stage II as thought by Van der Made (1996) whereas the canine attributed to Calydonius trux is rather too large to belong to this evolutionary stage, and more likely represents evolutionary stage III.
Metric analysis of the dentition. Comparisons of the dimensions of the cheek teeth of the Nebisuyu specimen with other listriodonts from Eurasia, indicates that they fall into the known range of variation of Listriodon splendens ( Tab. 1). In detail the specimen corresponds closely in dimensions with material often attributed to the subspecies Listriodon splendens major ROMAN, 1907 , or Listriodon splendens evolutionary stage III of Van der Made (1996) which ranges in age from the end of MN 7 to the beginning of MN 9. This subspecies was defined in the Iberian Peninsula (Aveiras de Baixo, Portugal) ( Roman 1907) and has been reported from several localities in Spain (Hernandez-Pacego and Dantin Cereceda 1915, Van der Made 1996, Pickford and Morales 2003) and may occur at the type locality of Listriodon splendens, La Chaux- de-Fonds, Switzerland, where it is known under the name Calydonius trux VON MEYER, 1846 .
Huge suid canine from Nebisuyu. The locality at Nebisuyu from which the large skull of Listriodon splendens was collected also yielded a left upper male canine in a fragment of maxilla (EUNHM PV 3075) ( Text-figs 10 View Text-fig , 11 View Text-fig ). The specimen is large within the context of Listriodon , and it is possible that it represents the same individual as the skull. Large canines of Listriodon splendens are known from St Quirze and Hostalets, Spain, Bonnefont ( Text-fig. 14 View Text-fig ) and La Grive, France, and La Chaux-de-Fonds, Switzerland, the last being the type specimen of Calydonius trux VON MEYER, 1846 ( Van der Made 1996). The specimen from St Quirze, in particular, has almost the same antero-posterior diameter as the Nebisuyu tooth (54 mm versus 55 mm for the Turkish specimen) ( Text-fig. 15 View Text-fig ).
The radicular part of the Nebisuyu canine is short, about 25 mm on the ventral side and ca. 73 mm dorsally where the root is covered by the canine flange. This measurement is of the same order of magnitude as that of the canine in the skull of Listriodon xinanensis CHEN, 1986 from China ( Chen 1986, Van der Made 1996).
Even though the Nebisuyu canine lacks the apex (broken off or worn away, so no enamel bands are preserved) what remains accords closely in morphology with a specimen of Listriodon splendens from Sinap , Turkey, but it is substantially larger. There is a prominent dorsal gutter backed by a ridge as in the specimen from Sinap ( EUNHM AS 92.644) (Textfig. 12) and the anterior gutter is weakly expressed. The almost planar wear facet caused by abrasion against the lower canine is similar in the two specimens, with a notched outer edge which makes a prominent step on the anterior surface of the tooth. The wear facet narrows towards the root, its inner end being close to the edge of the alveolus (21 mm in the Nebisuyu specimen, 14 mm in the Sinap tooth) .
Measured from the medial end to the outer notch, the wear facet is 64 mm long in the Sinap specimen and 112 mm in the specimen from Nebisuyu. This measurement corresponds to the length of that part of the lower canine that contacts the upper canine during thegosis ( Text-fig. 13 View Text-fig ).
Other unpublished material of the large variant of Listriodon splendens is known from the locality of Helsighausen, Switzerland ( Text-fig. 16 View Text-fig , Tab. 3). An upper central incisor and a right mandible with cheek teeth from fine sandstone deposits in the area falls into the upper part of the range of metric variation of the species ( Tab. 2).
Some unpublished specimens of large Listriodon splendens from Hostalets, Spain, curated in other institutions, are included in order to fill out the fossil record of the taxon ( Text-fig. 17 View Text-fig ).
Hernandez-Pacego and Dantin Cereceda (1915) published detailed descriptions of teeth, jaws and postcranial elements of Listriodon splendens from Cerro del Ortero, Spain, attributing the fossils to the large subspecies major. The authors also discussed and illustrated what they considered to be an unknown artiodactyl. Stehlin (in Hernandez-Pacego and Dantin Cereceda 1915: 137– 138, 260–261) in a written communication to the authors commented that, after examining casts of the latter specimens, they show affinities to Hyopotamus and more particularly to a specimen from Monte Massi, Italy, that he had identified as Anthracotherium ? meneghinii, but probably representing a new genus of anthracothere. In fact all the Spanish fossils concerned are juveniles of Listriodon splendens major and are not in any way related to anthracotheres. D i s c u s s i o n. The skull of Listriodon from Nebisuyu is of an old individual – the M3/s are heavily worn and all that remains of the M1/s are stubs with small remnants of enamel at the bases of the paracone and protocone. The premaxillae and parts of the nasals are broken off and the skull has suffered slight lateral compression and diverse fractures affect much of the surface of the specimen. Of the dentition, only the P2/–M3/ are preserved on both sides, but from the same locality there is a left upper canine in a fragment of maxilla which could well be from the same individual. The neurocranium is almost complete but has been slightly compressed from side to side and has suffered soft-sediment distortion and some displacement of bony parts. The sutures are difficult to make out due to the advanced age of the individual, but the contact between the lachrymal and frontal can be made out on both sides, the surface of the lachrymal being smooth, whereas that of the frontal above it is thickened and rugose.
What is remarkable about the Nebisuyu specimen is the fact that it preserves the frontals, which are rarely represented in the fossil record of the genus, the few specimens retaining this part of the skull being a large female of Listriodon splendens from Toril, Spain ( Pickford and Morales 2003), a fossil of Listriodon xinanensis from Xinan, China ( Chen, 1986) and a specimen from Bézian, France, attributed to Bunolistriodon lockharti ( POMEL, 1848) by Ginsburg and Bulot (1987). Lopholistriodon kidogosana from Ngorora, Kenya ( Pickford 1986) also preserves the frontals, but, as its name implies, it is a considerably smaller animal than Listriodon splendens . Specimens of Eurolistriodon adelli PICKFORD et MOYA- SOLA, 1995, from Els Casots, Spain and Eurolistriodon tenarezensis ORLIAC, 2006 , from Montréaldu-Gers, France preserve the frontals but the fossils are highly compressed and yield little information about their morphology. A partial skull from Chinji, Pakistan, also preserves part of the frontal bones ( Van der Made 1996). Thus, despite its crushed and distorted morphology, the Nebisuyu fossil is of great interest in revealing aspects of the morphology of part of the skull that is rarely represented in the palaeontological record.
Furthermore, the Nebisuyu skull is unusual among Suidae in showing thickened and inflated frontals which rise abruptly just distal to the naso-frontal zone, well in front of the orbits, thereby forming a solid base for a single sagittal horn-like protuberance, the ossicone. The apex of the ossicone itself is missing but its broken base measures about 50 mm antero-posteriorly by ca. 31 mm from side to side. Lower down, the solid base of the horn-like structure measures 65 mm across and it overhangs the lachrymals on both sides (7 mm on the left and 25 mm on the right, the latter measurement possibly exaggerated by inward, post-mortem displacement of the lachrymal). The swollen, inflated and somewhat rugose appearance of the surface of the frontals extends distally from this horn base and affects the post-orbital processes which are enlarged and show a rugose outer surface. Distal to the post-orbital processes the frontals display a more usual suid-like appearance of the fronto-parietal parts, the temporal ridges lying well apart from each other (i.e., no saggital crest is present). However, the nuchal area is deeply excavated and the distal margin of the temporo-nuchal crest invaginates anteriorly rather than being almost straight and transversely oriented as in many suids ( Stehlin 1899, 1900).
The only other suids that are known to have possessed frontal protuberances are Kubanochoerinae GABUNIA, 1958 ( Gabunia 1958, 1960) but the position and structure of the horns in this subfamily are different in the Nebisuyu specimen, the base of the horn being more distally positioned (above the orbits) and the frontal bones on which it is posed are not raised dorsally, nor are they thickened to any great extent. Some nyanzachoeres possess horn-like protuberances on the zygomatic arches ( Gebreyesus 2011, Reda et al. 2017) but these are radically different from the horns of listriodonts and kubanochoeres, not only by their positions, but also by the fact that they are present on both sides of the skull.













No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |