
Aspidoscopulia
|
publication ID |
https://doi.org/ 10.5281/zenodo.203661 |
|
DOI |
https://doi.org/10.5281/zenodo.5612034 |
|
persistent identifier |
https://treatment.plazi.org/id/345787A2-ED7C-FF80-3EF7-50DB1175FE27 |
|
treatment provided by |
Plazi |
|
scientific name |
Aspidoscopulia |
| status |
|
Aspidoscopulia sp.
( Figures 1 View FIGURE 1 , 12 View FIGURE 12 and 13 View FIGURE 13 )
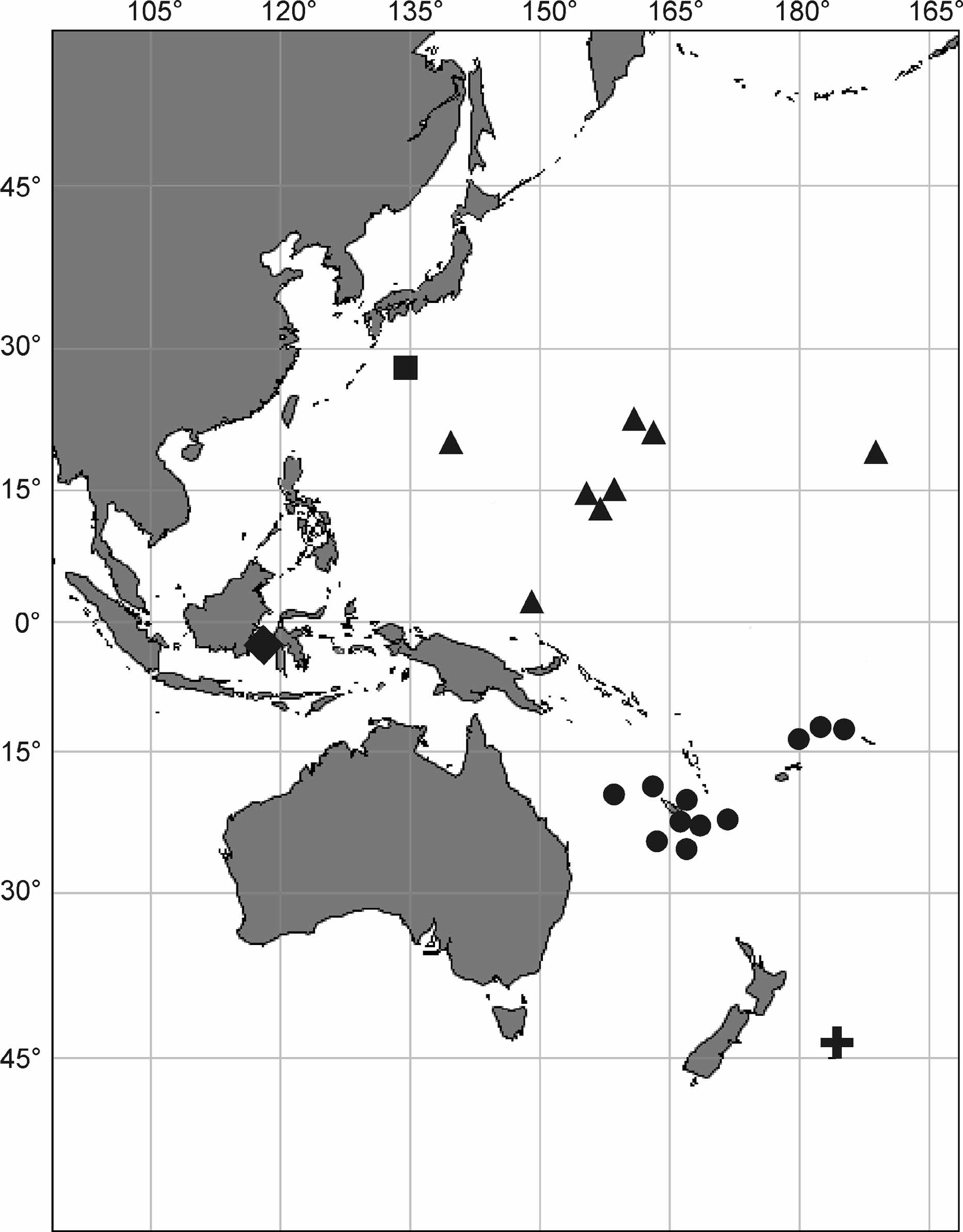
Material examined. Marcus-Necker Mountain chain ( Fig. 1 View FIGURE 1 ): R.V. ‘Vitjaz’ 48, stn. 6359.2, 19°02.8’N, 171°08.9’ W, 1270–1320 m: IORAS 5/2/3136. Stn. 6364, 21°10.0’ N, 163°13.2’ E, 2310–3085 m: IORAS 5/2/3137- IORAS 5/2/3159. Stn. 6366, 22°39.5’ N, 160°52.2’ E, 1900 m: IORAS 5/2/3199. Parese-Vela Basin ( Fig. 1 View FIGURE 1 ): R.V. ‘Academic Mstyslav Keldysh’ 9, stn. 1032, 20°14.3’–13.65’ N 139°51.0’– 50.7 E, 5139– 5132 m: IORAS 5/2/180. Magellan underwater Mountains (Ita-Maitai Guyot) ( Fig. 1 View FIGURE 1 ): R.V. ‘Academic Mstyslav Keldysh’ 9, stn. 1037, 12°51.7’– 51.2’ N, 157°01.8’– 01.6 E, 1620–2000 m: IORAS 5/2/107, IORAS 5/2/110, IORAS 5/2/111. Stn. 1042, 12°54.2’– 55.1’ N, 156°45.4’– 40.5 E, 1635–1728 m: IORAS 5/2/169. Stn. 1043, 12°54.39’ N, 156°42.48’ E, 1959 m: IORAS 5/2/172. Stn. 1044, 12°56.95’ – 55.5 N 156°46.05’–45.88’ E, 1600–1815 m: IORAS 5/2/160, IORAS 5/2/163, IORAS 5/2/194. Stn. 1047, 12°54.25’–54.2’ N, 156°49.1’– 47.9 E, 1450–1490 m: IORAS 5/2/119, IORAS 5/2/ 121,1. Stn. 1053, 12°58.25’–57.75’ N, 156°37.6’– 36.4 E, 4150– 4000 m: IORAS 5/2/173. Magellan underwater Mountains ( IOAN Guyot) ( Fig. 1 View FIGURE 1 ): R.V. ‘Academic Mstyslav Keldysh’ 9, stn. 1058, 14°13.0’ N, 155°57.5’– 58.8 E, 1530–2000 m: IORAS 5/2/113. Stn. 1059, 14°12.7’–12.85’ N, 155°58.15’–58.50’ E, 1485–2000 m: IORAS 5/2/0, IORAS 5/2/162. Stn. 1063, 14°10.2’ N 155°59.2’– 156°00.2’ E, 1410–2000 m: IORAS 5/2/134. Stn. 1065, 14°09.05’–08.5’ N, 155°54.4’–53.0’ E, 380–4270 m: 5/2/176, IORAS 5/2/178, 5/2/196. Stn. 1070, 14°08.94’– 09.7’ N, 156°31.95’–32.3’ E, 1500–1900 m: IORAS 5/2/116.1. Mussau Mauntain chain ( Fig. 1 View FIGURE 1 ): R.V. ‘Academic Mstyslav Keldysh’ 9, stn. 1074, 2°12.3’ N, 149°03.0’–03.44’ E, 1520–1930 m: IORAS 5/2/186, 5/2/189, 5/2/191, 5/2/202.1.
A. furcillata ( Levi, 1990) A. tetrasymmetrica sp. n. A. bisymmetrica sp. n.
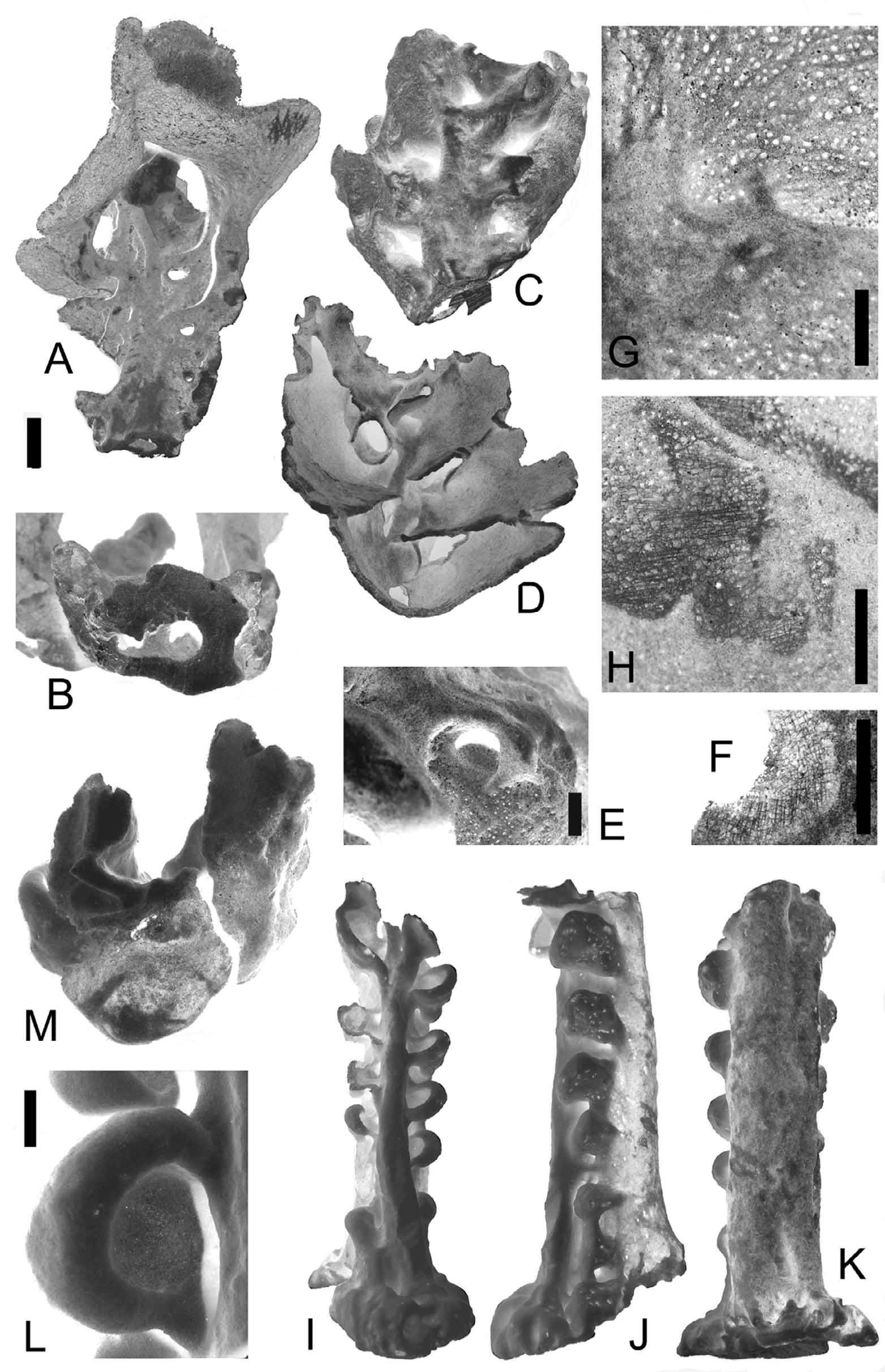
Description. Body: The size of some of these specimens is very large. As reconstructed from photos and collected fragments, the largest sponges may reach about 2 m in height and 1.5 x 0.5 m in the transverse section of the upper part. Other specimens are smaller – about 1.5 m high. The smallest specimen ( Fig. 13 View FIGURE 13 D) that is assigned with some hesitations to this genus is about 50–80 cm high. The measurements are obtained by indirect method after the comparison of collected and photographed specimens. All the collected specimens are remnants of large specimens, with construction type conforming mostly to that of the above described species, A. bisymmetrica sp. n. The construction of the upper parts of the specimens is more regular than that observed on the photo of A. aff. bisymmetrica ( Fig. 11 View FIGURE 11 ). The specimens ( Fig. 12 View FIGURE 12 , Fig. 13 View FIGURE 13 ) are considerably larger: the main stem of the lower part of the body is about 80 x 90 mm in section; the atrial cavity (about 10 mm in diameter in section) is overgrown by secondary framework, some lateral oscula are retained as cavities or depressions; the ear-like processes are regular, not fused with each other, 6–10 mm thick. The upper parts carry enlarged, meandering ear-like processes 4–6 mm thick, which fuse with their neighbors. The main stem is 30–40 mm in diameter, and the atrial cavity is 6–12 mm in diameter in section. The ear-like processes have numerous epirhyses (0.5–1 mm in diameter, 1–3 mm deep) on both sides, which penetrate the secondary framework and partly the primary one (when the meshes are large); the secondary framework of the main stem has no channelization, as well as atrial surfaces lined by the primary frameworks.
Dictyonal framework: The primary framework in the ear-like processes is represented by 6–8 (inner) layers; the number of layers of primary framework which underlies the atrial cavity is unknown in these large specimens because of the intensive growth of the secondary framework taking place inside the primary one, and atrial spaces are nearly entirely overgrown by the secondary framework. One may only suspect that the primary framework, which underlies atrial cavities, consists of 3–4 layers.
Remarks. It is likely that the discussed specimens belong also to Aspidoscopulia bisymmetrica sp. n., or to a close species, but absence of loose spicules in the collected material (all these sponges are dead fragments containing rigid dictyonal framework only) and considerable remoteness from the type location does not allow this problem to be solved now. The specimens make relatively sparse populations in the Magellan Mountains, likely inhabiting the investigated area for a very long time. The branching pattern observed on specimens from these locations resembles A. bisymmetrica sp. n. The observed differences are in size, in further developing of ear-like processes that become massive lobe-like, owergrowth of the atrial cavity and lateral oscula and finally in the absence of epirhyses on the main stem. This may be a result of extreme enlargement of the main stem. Thus, only the finding of smaller specimens with loose spicules in the same area may definitely settle the question of species identity.
The finding of the specimens in the living position was not very common, while some areas (it seems that ledges and their feet) accumulate great amounts of dead fragments of basal parts of these sponges covered by the iron-manganese crust. The analysis of numerous photos taken in this area with the ‘Pisces’ submersibles during the R.V. ‘Academic Mstyslav Keldysh’ 9 th expedition showed that young specimens of this sponge are very rare. The most reasonable explanation of this observation is that some factors working for a long time do not allow development of new, young specimens. Most probably the observed specimens are relicts of the distant past when different conditions promoted growth of these sponges. Another possibility is that the growth of these sponges is very slow and large accumulation of dead fragments represent very long time span under conditions of a very slow sedimentation rate.


| IOAN |
Shirshov Institute of Oceanography |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Aspidoscopulia
| Tabachnick, Konstantin R., Menshenina, Larisa L., Pisera, Andrzej & Ehrlich, Hermann 2011 |
A. furcillata (
| Levi 1990 |