
Aspidoscopulia bisymmetrica, Tabachnick, Konstantin R., Menshenina, Larisa L., Pisera, Andrzej & Ehrlich, Hermann, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.203661 |
|
DOI |
https://doi.org/10.5281/zenodo.5612028 |
|
persistent identifier |
https://treatment.plazi.org/id/345787A2-ED74-FF9F-3EF7-558F1408FBC0 |
|
treatment provided by |
Plazi |
|
scientific name |
Aspidoscopulia bisymmetrica |
| status |
sp. nov. |
Aspidoscopulia bisymmetrica sp. n.
( Figures 1 View FIGURE 1 , 5–10 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 ; Tables 2–3 View TABLE 2 View TABLE 3 )
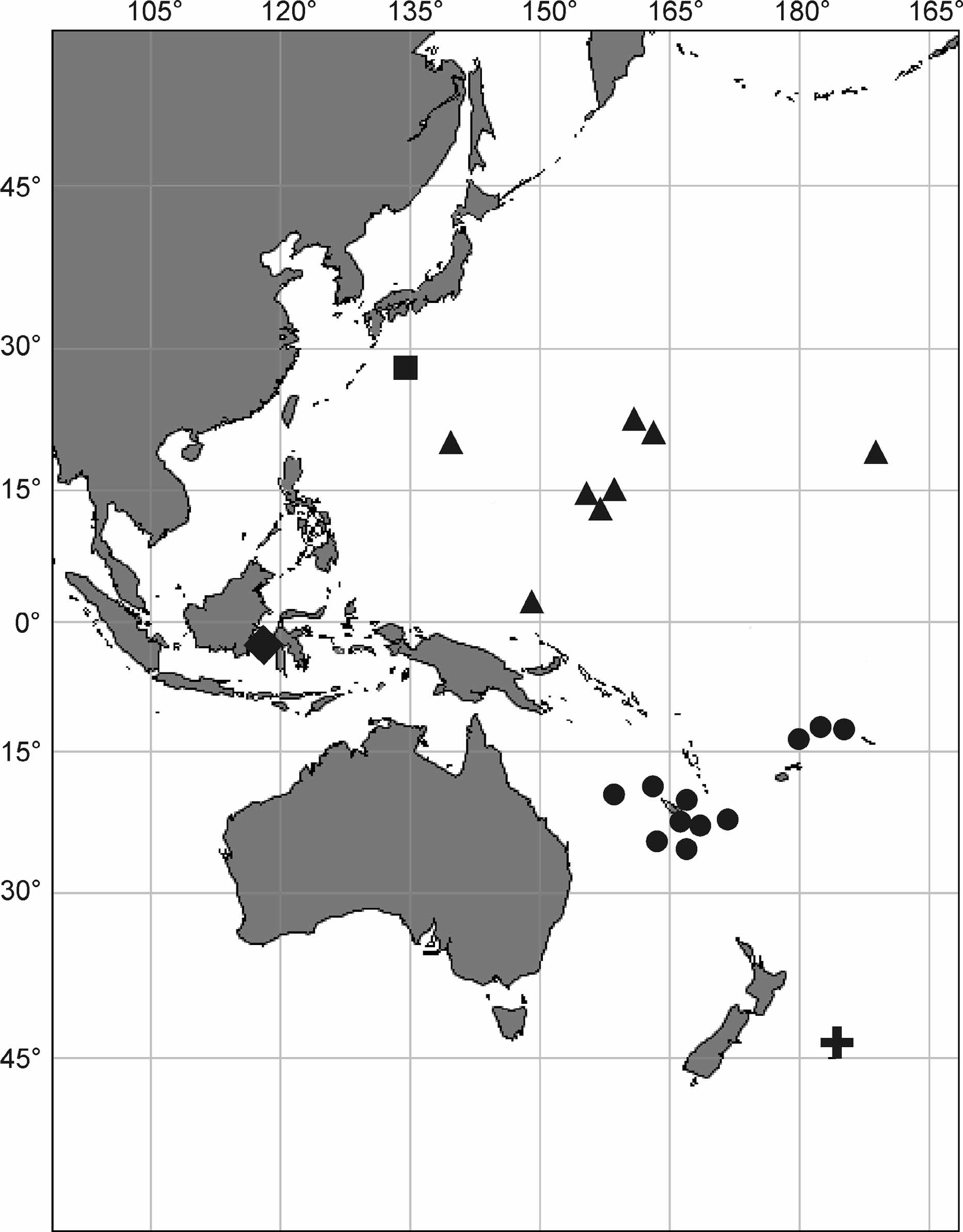
Holotype. MNHN fr546 ( Fig. 5 View FIGURE 5 C), Off Loyalty Islands ( Fig. 1 View FIGURE 1 ): Biogeocal, R.V. ‘Coriolis’, stn. DW 290, 20°36.91’ S, 167°3.34’ E, 920– 760 m.
Paratypes. MNHN fr534, 535, 536, 537, 538, 543, same to the holotype location. Off Loyalty Islands: Biogeocal, R.V. ‘Coriolis’, stn. CP 297, 20°38.64’ S 167°10.77’ E, 1230–1240 m: MNHN fr494. Musorstom 6, R.V. ‘Alis’, stn. CP 466, 21°5.25’ S, 167°32.2’ E, 540 m: MNHN p1219. Stn. DW 488, 20°49.2’ S 167°6.44’ E, 800 m: MNHN p3701. Off New Caledonia ( Fig. 1 View FIGURE 1 ): Biocal, R.V. ‘Jean Charcot’, stn. DW 80, 20°31.69’–31.86’ S, 166°48.35’–48.59’ E, 900–980 m: MNHN p61. Halipro - 2, R.V. ‘Zoneco’, stn. BT 0 52, 25°21.45’ S, 168°16.94’ E, 810–1172 m: MNHN p5019. Stn. BT 0 63, 24°39.72’ S 168°41.82’ E, 782–1100 m: MNHN p5020. Musorstom 4, R.V. ‘Vauban’, stn. CP 199, 18°50’ S, 163°14.5’ E, 600 m: MNHN p3744. Volsmar, R.V. ‘Alice’, stn. DW 4, 22°24.7’–22.4’ S, 171°49’–49.1’ E, 825–850 m: MNHN p3732, p3733, p3734, p3735. Off Wallis and Futuna Islands ( Fig. 1 View FIGURE 1 ): Musorstom 7, stn. CP 551, 12°15.3’ S, 177°28.1’ W, 791–795 m: MNHN p3674. Stn. CP 592, 12°32.4’ S, 174°22’ W, 775– 730 m: MNHN p1142. Stn. DW 637, 13°37’ S, 179°56’ W, 820–830 m: MNHN p6123. Norfolk Ridge ( Fig. 1 View FIGURE 1 ): Norfolk II, stn. 2053, 23.661° S, 168.260° E, 67–708 m: ZPAL Pf.22/wa75. Stn. 2054, 23.660° S 168.253° E, 736–800 m: ZPAL Pf.22/wa115, wa117. Stn. 2055, 23.654° S 168.274° E, 900–950 m: ZPAL Pf.22/wa70, wa71, wa72, wa73. Stn. 2065, 25.261° S 168.927° E, 750–800 m: ZPAL Pf.22/wa86.
Etymology. The name refers to the bilateral symmetry of lateral branches, which arise from the main stem of the sponge body.
Diagnosis. Aspidoscopulia with two rayed symmetry as seen from the top; clavules mostly with discoidal-clavate (pileate), and some small with anchorate heads; microscleres with oxyoidal, discoidal and onychoidal outer ends.
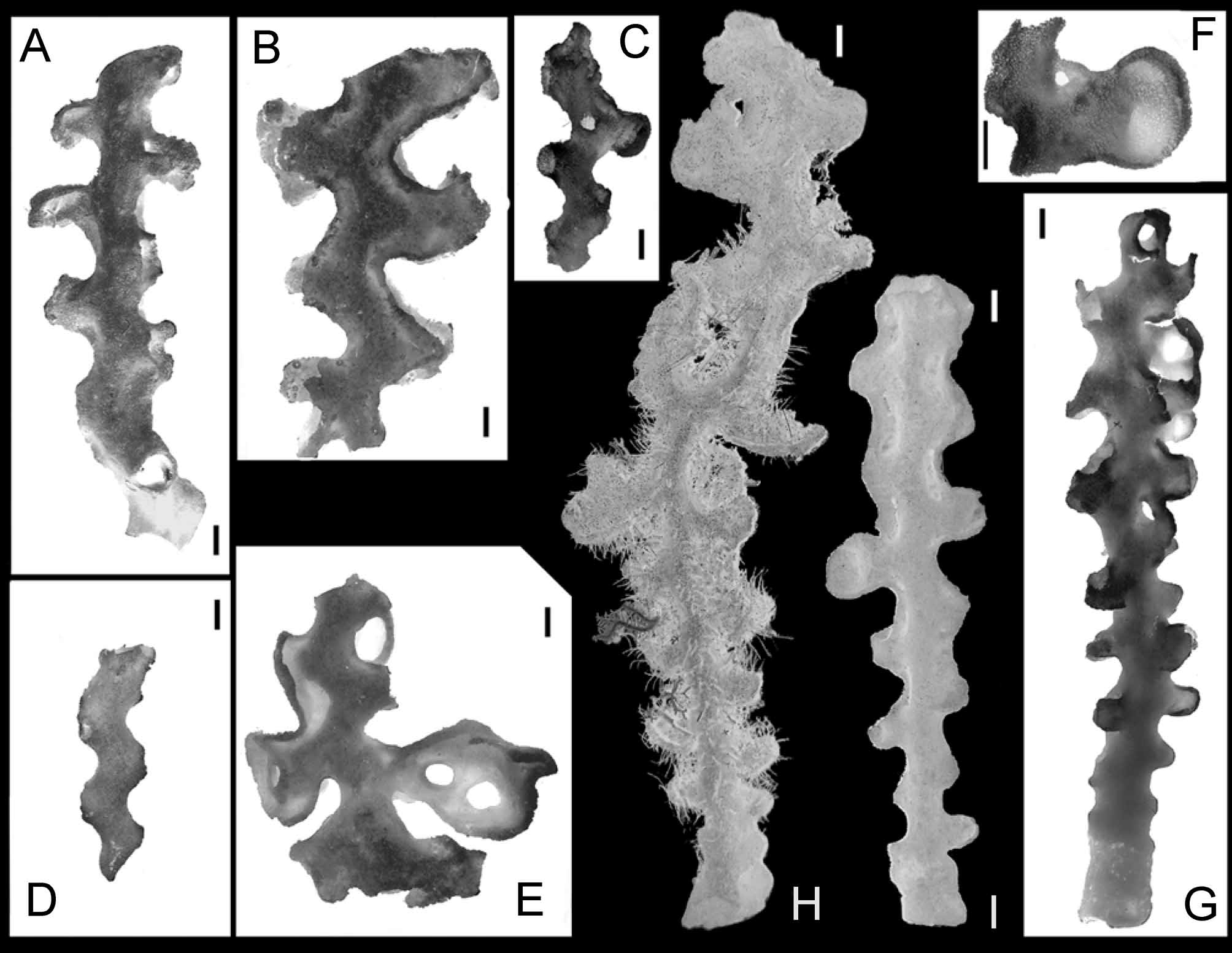
Description. Body: Sponges are mostly represented by the main zigzag-shaped stem with lateral branches ( Fig. 5 View FIGURE 5 ). The lateral branches are situated along the main stem and consist of very short unbranched tubes or, rarely, dichotomously branching into two short secondary branches. The neighboring tubes or openings of lateral oscula and other derived constructions are oriented at right angle to the main stem. The lateral structures (branches, lateral oscula and derived constructions) are situated in regular, alternate position on the main stem in two opposite rows. In result, a 2-rayed symmetry is observed in these sponges. Common structures are ridges (usually two parallel, sometimes one) along the main stem connecting the bases of lateral branches ( Fig. 5 View FIGURE 5 B). These ridges are variously developed, from low and hardly distinguishable to relatively high with occasional apertures. The ear-like processes ( Fig. 5 View FIGURE 5 F) are developed by side-by-side wall fusion and further growing up and enlargement of the space between two secondary lateral oscula ( Fig. 6 View FIGURE 6 e-lp, Fig. 7 View FIGURE 7 C). Sometimes, usually in the lower part of the body, the same branching process leads to common formation of the carina (line of fusion of the walls of lateral branches) and equal dichotomous division of the lateral tube ( Fig. 5 View FIGURE 5 A, Fig. 6 View FIGURE 6 lo, dblb, Fig. 7 View FIGURE 7 B). This results in the appearance of two secondary lateral branches (equal to each other and to their stem) with two secondary lateral oscula. The secondary lateral oscula which accompany the ear-like processes are smaller in diameter than their homologues in dichotomous-isotomous branching (here and bellow we use terminology of plants branching); besides they are often completely overgrown with secondary skeletal framework ( Fig. 6 View FIGURE 6 slo). The ear-like processes are always becoming spoon-like with uniform orientation; usually they fuse with their further neighbors in the upper part of the sponge, as may be observed in some fragments ( Fig. 5 View FIGURE 5 E), they somehow make irregular structures of curved and anastomosing lamellae. In the terms of branching, this species in whole is anisotomous (in small specimens) with tendency to dichopodial-monopodial branching (in large specimens). Unfortunately no complete specimen was ever found in the collections. The living specimens may be sometimes covered by dense aggregations of hydrozoan and zoantharian epibionts.
The holotype ( Fig. 5 View FIGURE 5 C) is a fragment of the main stem 75 mm in length consisting of a main tube 8–12 mm in diameter with walls 1–2 mm thick. Primary lateral oscula are 8–14 mm in diameter, the ridges between them are low, up to 3 mm high and 1 mm in thickness. One large, flat, ear-like process vertically oriented, protrudes at about 12 mm and is about 2 mm thick. It has two small secondary lateral oscula about 3 mm in diameter, situated at base and on the upper part of the ear-like processes. The paratypes are different fragments in which the stem tube and its outgrowths are 8–17 mm in diameter, sometimes with some ear-like processes up to 40 mm in diameter.
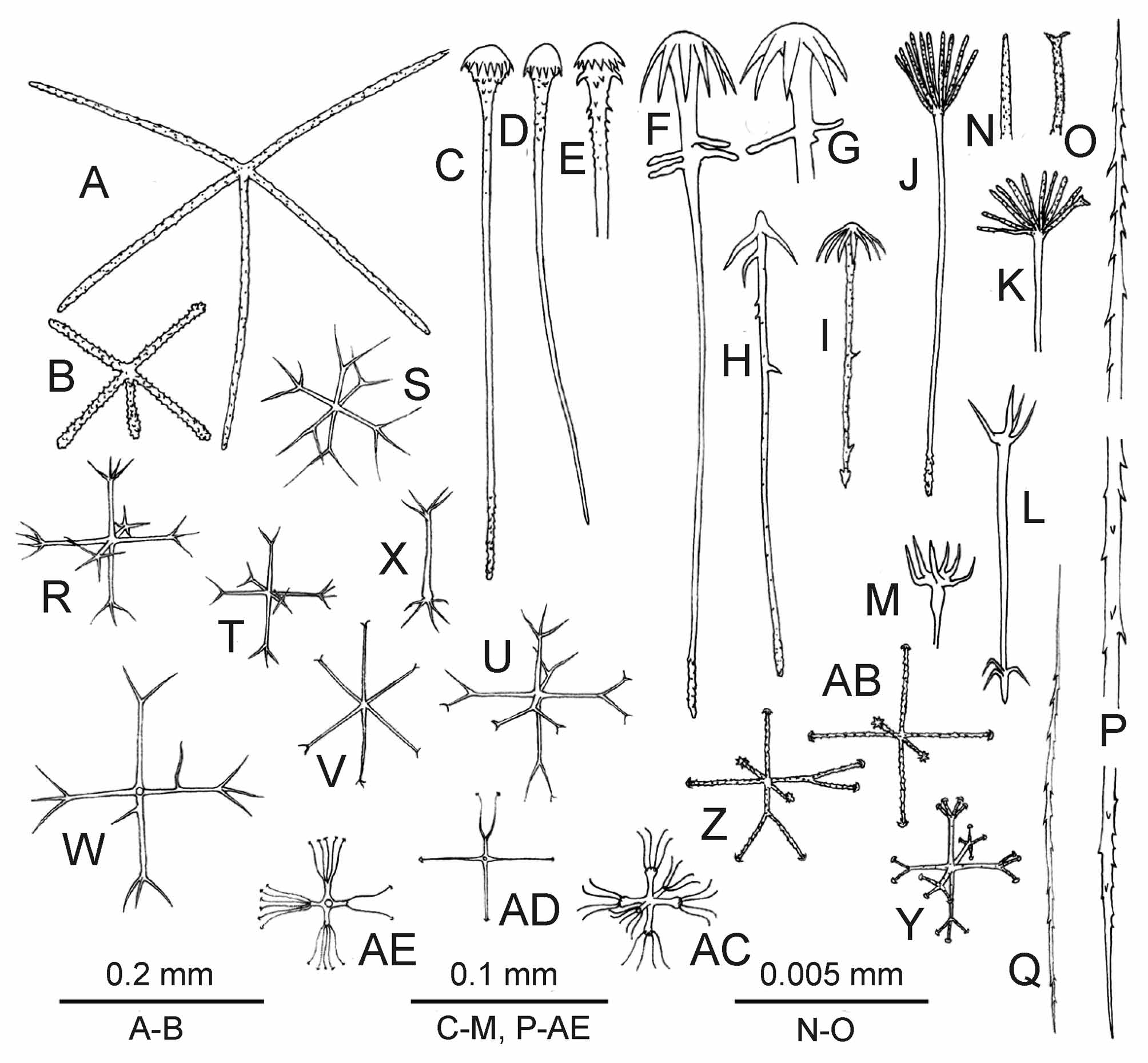
Dictyonal framework: Dictyonal skeleton of euretoid type. Most tubes have inner walls of 3 layers of primary skeleton constructed of smooth beams 40–110 μm in diameter and rectangular meshes about 200x500 μm in size, with free rays that are rough and about 0.5 mm long. The primary skeleton ( Fig. 8 View FIGURE 8 A, B) underlies not only the atrial cavity but also it is present as several (4–6) inner layers of the ear-like processes (the secondary skeleton in the earlike processes has 2–3 layers on both sides). The secondary skeleton ( Fig. 8 View FIGURE 8 C) in the upper parts of the body is constructed by beams 20–80 μm in diameter with usually triangular meshes 50–200 μm in diameter. The small hexactines connected by fusion with dictyonal skeleton and with each other have smooth rays 50–110/4–8 μm in size ( Fig. 8 View FIGURE 8 D). Epirhyses (100–200 μm in diameter, up to 500 μm deep) are connected with all secondary framework structures; they are situated on both sides of the ear-like processes and on the surface of the main stem and lateral branches.
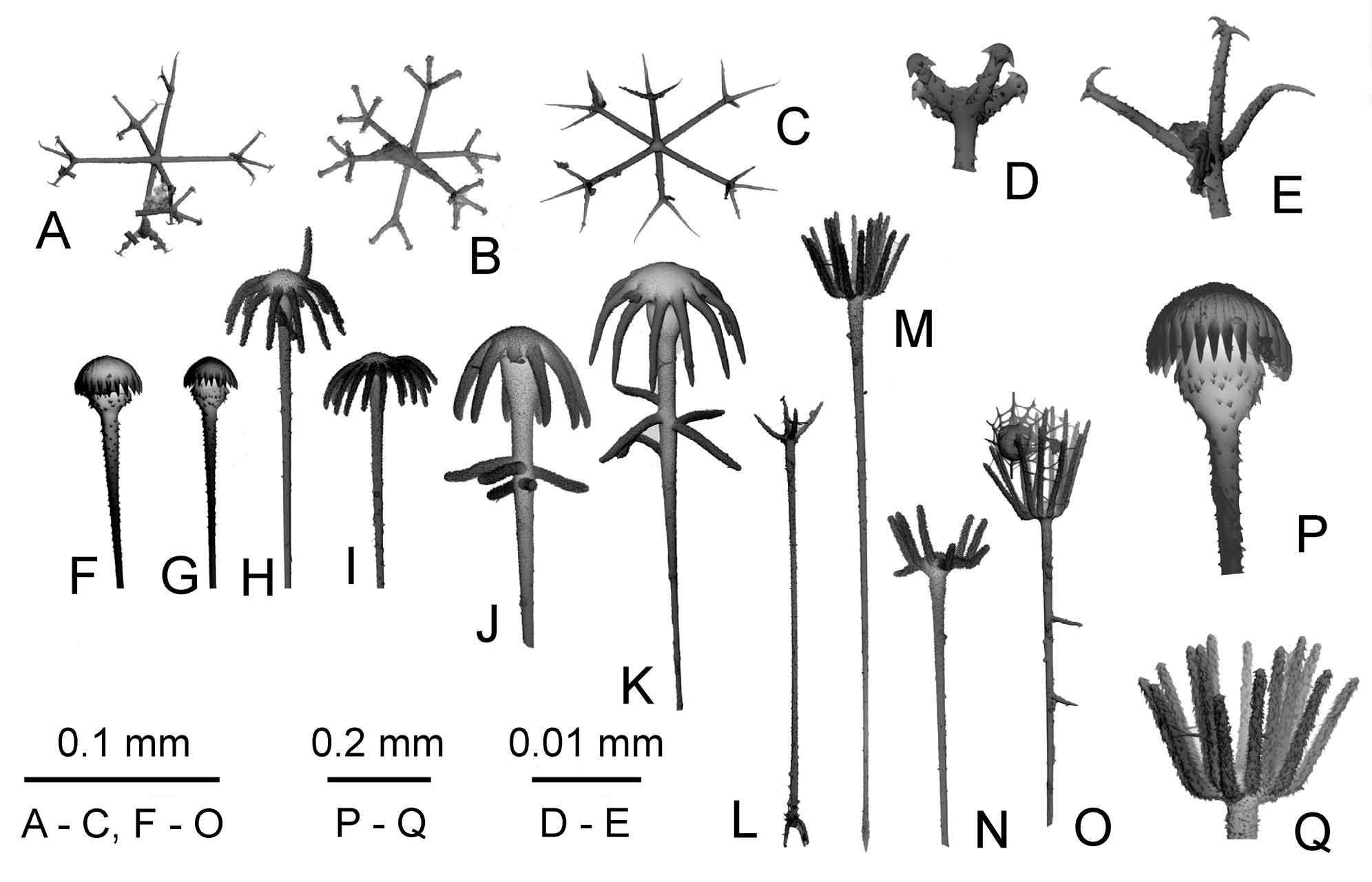
Free spicules: Dermalia and atrialia are pentactines ( Fig. 10 View FIGURE 10 A, B) with unpaired ray directed inside the body. Dermal pentactines are usually larger than atrial ones, with smoother rays and with less clavate outer ends. The dermal pentactines are generally larger and their ray surface is smoother than in artrial ones. The tangential rays of dermal pentactines are 107–366 μm long, the unpaired ray is 67–348 μm long, and 7–11 μm in the diameter. The tangential rays of atrial pentactines are 59–307 μm long, the unpaired ray is 44–340 μm long and 4–9 μm in the diameter. Clavules are of two types: most have discoidal-clavate (pileate) ( Fig. 9 View FIGURE 9 F, G, P, Fig. 10 View FIGURE 10 C–E), and some anchorate heads (8–9 teeth) ( Fig. 9 View FIGURE 9 J, K, Fig. 10 View FIGURE 10 F–I), the latter have several spines, which are long, curved, and situated close to the head (in some specimens the latter type of clavules was not found). Their shafts are slightly rough; the end directed inside the body is rough-spiny, usually lanceolate in shape. The discoidal clavules are 196– 344 μm long, their heads are 11–26 μm long and 19–44 μm in diameter, the diameter of the shaft is about 2 μm in the middle. The anchorate clavules are 133–426 μm long, their heads are 19–56 μm long and 33–63 μm in diameter, the diameter of the shaft is about 2 μm in the middle. The aspidoscopules ( Fig. 9 View FIGURE 9 L– O, Q, Fig. 10 View FIGURE 10 J–M), are found only in the primary framework which underlies the atrial space (they are absent in the primary framework inside the ear-like processes). The aspidoscopules have 12–14 terminal spines with conically pointed or rarely dichotomously branching outer ends and microspined surface. Their shafts are similar in shape to that of the anchorate clavules. The aspidoscopules are 118–352 μm long, their heads are 7–56 μm long and the tuft is 17–70 μm in diameter. The diameter of the shaft is about 3 μm in the middle. The clavules are situated at dermal surface while the aspidoscopules at atrial one. Uncinates ( Fig. 10 View FIGURE 10 P) are 200–1000/2–8 μm in dimensions.
Microscleres: Oxyhexasters ( Fig. 9 View FIGURE 9 C, Fig. 10 View FIGURE 10 R–T), onychohexasters ( Fig. 9 View FIGURE 9 A, E, Fig. 10 View FIGURE 10 U) and discohexasters ( Fig. 9 View FIGURE 9 B, E, Fig. 10 View FIGURE 10 Y) with 2–4 – rarely 5 – secondary rays, are 50–137 μm in diameter with primary rosette 25–81 μm in diameter. Rare hexactines and hemihexasters (1–4 secondary rays) are found in some specimens, they have onychoidal–discoidal outer ends and diameter 54–101 μm ( Fig. 10 View FIGURE 10 U-AD). These spicules may be allochthonous. The holotype and additional topotypical specimens (may be its fragments) have numerous small stellate discohexasters ( Fig. 10 View FIGURE 10 AC) 25–54 μm in diameter with primary rosette 7–36 μm in diameter. Such spicules were not found in other paratypes taken from other locations, and one might conclude that they are allochthonous. This idea is rejected here because they are very numerous in comparison with other definitely allochthonous spicules. Abnormal microscleres are rare oxyhexasters with very short secondary rays ( Fig. 10 View FIGURE 10 U, V) (MNHN p3701), oxydiasters ( Fig. 10 View FIGURE 10 X) (MNHN p5019, p3735; fr542), and oxystaurasters (MNHN p1142; fr543).
Remarks. The three known species of Aspidoscopulia differ in the following characters ( Table 3 View TABLE 3 ): A. furcillata ( Lévi, 1990) has no anchorate clavules and microscleres with short primary rays; A. tetrasymmetrica sp. n. has large anchorate clavules (200–600 μm long) and only oxyoidal microscleres, while A. bisymmetrica sp. n. has small anchorate clavules (130- 140 μm long) and microscleres with oxyoidal, discoidal and onychoidal outer ends. Besides these features the species differ in their external shape (see the descriptions) but that of A. furcillata is unknown – this sponge is represented by a fragment for which body shape interpretation is impossible. Many specimens of ‘bilateral shape’ collected far from A. bisymmetrica sp. n. have no loose spicules. Nevertheless it is impossible to assign a specific body form to a particular species now. A very possible representative of this species is shown on the underwater photo ( Fig. 11 View FIGURE 11 ) taken near the type locality.
continued.
MNHN fr534 MNHN p3701
n avg min max std n avg min max std continued.
MNHN p3735 MNHN p5019



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.